
فبركة جديدة من موقع العربية نت بعد فبركة “الحوت الأزرق”:
بسيكولوجيا
الدماغ البشري (15)
التحديق المزدوج conjugate gaze يعني تحريك العينين في آن واحد نحو نفس الهدف. الخلل في هذه العملية يضعف جودة الرؤية ثنائية العين binocular vision ويسبب الرؤية المزدوجة diplopia. التحديق المزدوج يمكن أن يكون في ثلاثة اتجاهات أساسية:
- التحديق العمودي vertical gaze (تحريك العينين إلى الأعلى أو الأسفل)
- التحديق الجانبي أو الأفقي lateral or horizontal gaze (تحريك العينين إلى اليمين أو اليسار)
- المقاربةconvergence (تحريك العينين إلى الداخل)
 الطَّرْف والتثبيت والتعقب السلس
الطَّرْف والتثبيت والتعقب السلس
الطَّرْف saccade هو أحد أساليب تحريك العينين عند البشر. العينان تتحركان بهذا الأسلوب للنظر نحو جسم جديد دخل إلى مجال الرؤية.
أعضاء الحس عند البشر تتعامل في كل لحظة مع عدد هائل من المؤثرات الحسية، ولكن كثيرا من هذه المؤثرات الحسية هي قليلة الأهمية. على سبيل المثال، عندما أستمع إلى شخص يتحدث عبر مكبر صوت فإنني ربما أسمع ضجيجا مرافقا لكلامه، ولكن هذا الضجيج هو ناتج من الآلة وليس من كلام الرجل، وهو غير مهم بالنسبة لي مقارنة بالكلام الذي يقوله الرجل.
عندما أشاهد التلفاز فإنني ربما أشاهد بعض التشويش على الشاشة، ولكن هذا التشويش هو غير مهم بالنسبة لي مقارنة بالأمور الأخرى التي تظهر على الشاشة.
كثير من الأحاسيس التي تصل إلى الدماغ هي مجرد تشويش أو أحاسيس قليلة الأهمية. لهذا السبب الدماغ لا يستطيع أن يتعامل بجدية مع كل الأحاسيس التي تصل إليه وإلا فإنه سيصاب بالضياع والاستنزاف.
الدماغ يصنف الأحاسيس حسب أهميتها. الدماغ يعتبر أن بعض الأحاسيس التي تصل إليه هي أهم من غيرها، وبناء على ذلك هو يقوم بتوجيه طاقته الإدراكية نحو هذه الأحاسيس المهمة. هذه العملية تسمى “الانتباه” attention. المقصود بالانتباه هو توجيه طاقة الإدراك الحسي نحو أحاسيس معينة على حساب أحاسيس أخرى.
المؤثرات الحسية التي تجذب انتباه الدماغ أكثر من سواها تسمى مؤثرات حسية “بارزة” salient. بروز المؤثر الحسي salience يعتمد على عدة معايير. من هذه المعايير اختلاف المؤثر الحسي عن المؤثرات الأخرى المرافقة له. مثلا لو شاهدت ورقة بيضاء فيها 49 نقطة خضراء ونقطة واحدة حمراء فإن النقطة الحمراء ستجذب انتباهي، أي أنها ستكون بارزة salient. أيضا ندرة المؤثر الحسي لها دور في بروزه. مثلا لو كنت أستمع طوال اليوم إلى نغمة معينة ثم سمعت فجأة نغمة مختلفة فإن النغمة المختلفة ستجذب انتباهي.
الذاكرة أيضا تلعب دورا في تحديد بروز المؤثر الحسي. الإنسان يختزن في ذاكرته أمورا معينة تعتبر بالنسبة له أمورا مهمة، وعندما يستقبل مؤثرات حسية لها علاقة بهذه الأمور المختزنة في ذاكرته فإنه يتعامل معها بانتباه.
حركة الطَّرْف saccade هي الطريقة التي تتعامل بها العينان مع المؤثرات البصرية البارزة salient التي تدخل فجأة إلى مجال الرؤية. الطَّرْف يعني تحريك العينين بأقصى سرعة ممكنة نحو نقطة معينة في مجال الرؤية. مثلا لو كنت أراقب السماء ثم ظهرت فجأة طائرة في السماء فإن عيناي ستنفذان حركة طَرْف نحو موقع الطائرة. بعد ذلك سيقوم دماغي بتثبيت عيني على الطائرة لكي يتمكن من إدراكها. عملية التثبيت fixation تتطلب تدخل الأنوية الدهليزية vestibular nuclei في جذع الدماغ بالتعاون مع المخيخ cerebellum.
الطائرة لن تتوقف عندما أثبت عيني عليها ولكنها ستستمر في التحرك. لهذا السبب سيقوم دماغي بتنشيط آلية تحديق خاصة تسمى “التعقب السلس” smooth pursuit. ما يحدث في هذه الآلية هو أن عيناي تتحركان بنفس السرعة التي تتحرك بها الطائرة في مجال الرؤية بهدف إبقاء صورتها ثابتة على نقرة الشبكية fovea.
المؤثرات البارزة المعقدة
في الأعلى أنا ضربت أمثلة على مؤثرات حسية بارزة ذات تعقيد بسيط نسبيا (مثلا نقطة حمراء إلخ)، ولكن كثيرا من المؤثرات الحسية البارزة التي يتعامل معها الدماغ هي ذات تكوين معقد.
على سبيل المثال، الإنسان يرى في حياته اليومية الكثير من الوجوه faces. بعض الوجوه التي يراها الإنسان في حياته اليومية هي ليست بارزة (مثلا وجوه الناس المارين في الشارع)، ولكن هناك وجوها أخرى يهم الإنسان أن يتعرف على أصحابها، ولكي يتعرف على أصحابها لا بد له أن ينتبه إليها ويدقق فيها.
التمييز بين الوجوه المختلفة هو أمر صعب، لأن وجوه البشر تشبه بعضها، والإنسان يجب أن يدقق في الكثير من تفاصيل الوجه حتى يتمكن من تمييزه عن غيره. هو طبعا يجب أن يحفظ تفاصيل الوجه في ذاكرته إن أراد التعرف عليه لاحقا. من الأمور اللافتة التي كشفتها الأبحاث الحديثة أن هناك منطقة كبيرة في قشرة المخ متخصصة في حفظ الوجوه. هذه المنطقة تقع في أسفل الفص الصدغي temporal lobe على الجانب الوحشي للتلفيف المغزلي fusiform gyrus وتسمى أحيانا “منطقة الوجوه المغزلية” fusiform face area.

عندما يرى الإنسان وجها بارزا salient فإن عينيه تنفذان عددا كبيرا من حركات الطرف بهدف التعرف على الوجه:

الرسم أعلاه يبين النقاط التي تنظر إليها العينان بهدف التعرف على وجه. من هذا الرسم يتبين أن العينين تركزان على محيط الوجه والعينين والأنف والفم.
الدماغ عندما يرى جسما معقدا (كالوجه) فإنه يركز في البداية على دراسة النقاط الأهم في هذا الجسم، لأن دراسة هذه النقاط تساعد الدماغ على التعرف على ماهية الجسم في أسرع وقت ممكن.
الأبحاث الحديثة كشفت إلى حد ما آلية الإدراك البصري visual perception في الدماغ البشري. من الأمور اللافتة التي كشفتها الأبحاث أن الدماغ يقوم بتجريد الأجسام المرئية إلى عناصر بسيطة شبيهة بـ “الكاتغوريات” categories التي تحدث عنها الفيلسوف Immanuel Kant. الدماغ عندما يرى جسما فإنه في البداية يحلل هذا الجسم وفق الكاتغوريات التالية:
- مواقع النقاط المكونة للجسم وحدودها (الشكل الهندسي للجسم)
- ألوان النقاط المكونة للجسم (لون الجسم)
- اتجاه حركة النقاط المكونة للجسم (اتجاه حركة الجسم)
هذا التحليل المبدئي يتم في شبكية العين retina ويتكرر مجددا في الجسم الركبي الوحشي lateral geniculate body والتليلتين العلويتين superior colliculi والقشرة البصرية visual cortex. بعد هذا التحليل المبدئي يقوم المخ بمعالجة أكثر عمقا للجسم المرئي وفقا للمعلومات المخزنة في الذاكرة البصرية في الفص الصدغي temporal lobe ووفقا للمعالجة المنطقية في القشرة أمام الجبهية prefrontal cortex. القشرة الجدارية parietal cortex تقوم بإدراج الجسم المرئي في الخارطة الفراغية للعالم المحيط. (الخطوة الأخيرة تشبه كلام Kant عندما قال أن الفراغ والزمن space and time هما مفهومان غريزيان intuitive مزروعان في عقل الإنسان وليسا من المدركات الحسية.)
طريقة الإدراك الحسي في الدماغ البشري تتفق مع النظرية التي طرحها الفيلسوف Immanuel Kant والتي تسمى “المثالية العبارة للحدود” transcendental idealism. أنا شرحت هذه النظرية سابقا بشكل مختصر، ولدي النية لكي أعود لها في المستقبل، ولكنني الآن أردت فقط أن أشير إلى الأسلوب الذي يتعامل به الدماغ مع الأجسام المرئية. هذا الأسلوب هو بعيد عن أسلوب “الرسم النقطي” raster graphics or bitmap graphics الذي تعتمد عليه الكثير من برامج الكومبيوتر.
في أسلوب “الرسم النقطي” تُقسَم الصورة إلى عدد كبير من النقاط الصغيرة التي تسمى بكسلات pixels. لكي يدرك برنامج الكومبيوتر الصورة لا بد له أن يدرك جميع بكسلاتها. كل البكسلات متساوية في نظر البرنامج ولا توجد أهمية لبكسل على آخر.
أسلوب “الرسم النقطي” يتميز بأنه سهل من الناحية البرمجية، بمعنى أنه لا يتطلب برمجة معقدة، ولكن مشكلته هي أنه يستنزف الكثير من مساحة التخزين وطاقة المعالجة.
هناك أسلوب آخر في الرسم الكومبيوتري يسمى vector graphics. في هذا الأسلوب تُحلَّل الصورة إلى عناصر تجريدية على نحو شبيه نوعا ما بما يحصل في الدماغ البشري.

الصور المرسومة بطريقة vector graphics هي ليست مقسمة إلى بكسلات وإنما تتكون من عناصر هندسية. مثلا لكي نرسم دائرة بهذه الطريقة يجب أن ندخل معادلة الدائرة وندخل معها الإحداثيات الهندسية. الدائرة الناتجة يمكن أن تكون كبيرة المساحة على الشاشة، ولكن تخزينها في الكومبيوتر لا يتطلب سوى تخزين سطر واحد هو معادلة الدائرة. لو أردنا أن نرسم نفس الدائرة بطريقة البكسلات فسنضطر لتخزين المعلومات الخاصة بكل بكسل من بكسلات الصورة، وهو ما يتطلب عددا كبيرا جدا من الأسطر.
الرسم بطريقة vector graphics يمسي صعبا إذا كانت الصورة معقدة وتحوي الكثير من التفاصيل الدقيقة، ولكن ميزة هذه الطريقة في الرسم هي قلة مساحة التخزين وطاقة المعالجة المطلوبة مقارنة بالطريقة الأخرى.
الدماغ البشري يحلل المدركات المرئية بطريقة شبيهة بالـ vector graphics، وهذا يسهل على الدماغ معالجة المدركات المرئية وتخزينها. بالإضافة إلى ذلك الدماغ يملك القدرة على تصنيف العناصر المكونة للمدركات وفقا لأهميتها. مثلا الدماغ عندما يتعامل مع الوجوه يركز في البداية على العناصر الأكثر أهمية فيها (المحيط، العينان، الأنف، الفم)، ولهذا السبب نجد أن العينين عندما تريان وجها فإنهما تقومان بسلسلة من حركات الطرف saccades الموجهة نحو مناطق محددة في الوجه.
باحثو الجهاز العصبي يطلقون مسمى feature detection (“رصد الخصائص”) على عملية تحليل العناصر المكونة للمؤثر الحسي وتصنيف هذه العناصر حسب أهميتها.
آلية التحديق

تحريك العين الواحدة يتم عبر الأنوية القحفية رقم 3 و4 و6. النواتان القحفيتان رقم 3 و4 (الموجودتان في الدماغ الأوسط) تحركان العين في كل الاتجاهات ما عدا الاتجاه الخارجي أو الوحشي lateral. النواة القحفية رقم 6 (الموجودة في الجسر pons) تحرك العين في الاتجاه الوحشي عبر تقليص العضلة المستقيمة الوحشية lateral rectus muscle.
التحديق المزدوج conjugate gaze يتطلب ربطا عصبيا بين الأنوية المحركة لكل من العينين. هذا الربط يتم عبر أنوية وطرق عصبية في جذع الدماغ.

التحديق العمودي vertical gaze والمقاربةconvergence لا يتطلبان تحريك العين نحو الجهة الوحشية lateral، ولهذا السبب هما لا يتطلبان تدخل العصب القحفي رقم 6 (العصب المبعد abducens).
تنسيق التحديق العمودي والمقاربة يتم عبر أنوية موجودة في التشكل الشبكي للدماغ الأوسط midbrain reticular formation بالاشتراك مع النواتين القحفيتين رقم 3 و4.
تنسيق حركة المقاربة يتم عبر نواة تسمى “النواة فوق المحركة العينية” supraoculomotor nucleus. هذه النواة توجد على الجهة الظهرية للنواة المحركة العينية oculomotor nucleus.
تنسيق حركة التحديق العمودي يتم عبر نواتين توجدان على الجهة المنقارية للنواة المحركة العينية هما:
- Rostral interstitial nucleus of medial longitudinal fasciculus (riMLF)
- Interstitial nucleus of Cajal (INC)
الرسم التالي يوضح موقع هاتين النواتين. هذا الرسم هو لمقطع سهمي sagittal في جذع دماغ قرد:

على الرسم أعلاه تظهر النواتان INC و riMLF على الجهة المنقارية لنواة العصب القحفي الثالث III. هناك نواة ثالثة تظهر بجوارهما هي (ND) nucleus of Darkschewitsch. هذه النواة تسمى أيضا “نواة الملتقى الخلفي” nucleus of the posterior commissure لأنها تقع على الجهة الذيلية للملتقى الخلفي وهي مصدر معظم الألياف المكونة لهذا الملتقى. لا أعلم ما هي وظيفة هذه النواة، ولكن موقعها يوحي بأن لها علاقة بحس البصر.
حركة التحديق الجانبيlateral gaze تتطلب تحريك إحدى العينين نحو الجهة الوحشية، لهذا السبب هذه الحركة تتطلب تدخل نواة العصب القحفي رقم 6 الموجودة في الجسرpons . النواة الشبكية المسؤولة عن تنسيق حركة التحديق الجانبي هي النواة الشبكية جانب المحورية paramedian reticular nucleus، وتحديدا قسمها الجسري الذي يسمى(PPRF) paramedian pontine reticular formation .


النواة الشبكية جانب المحورية هي إحدى الأنوية الشبكية “قبل المخيخية” precerebellar (الأنوية الشبكية التي ترسل أليافها نحو المخيخ). هذه النواة تتلقى أليافا من نواة المخيخ الأَوْجِية fastigial nucleus التي أظن أنها مسؤولة عن تنسيق تحريك القسم المركزي median أو المحوري axial من الجسم (الجذع والرأس ومفاصل الأطراف القريبة من الجذع proximal دون المفاصل البعيدة عن الجذع distal).
القسم الجسري (المنقاري) من النواة الشبكية جانب المحورية PPRF هو متصل مع الأنوية القحفية المحركة للعينين بهدف تنسيق حركة التحديق الجانبيlateral gaze . هذا القسم يسمى “مركز التحديق الجانبي” lateral gaze center أو “مركز التحديق الأفقي” horizontal gaze center أو “مركز التحديق الجسري” pontine gaze center. بالنسبة للقسم النخاعي (الذيلي) من النواة الشبكية جانب المحورية فهو على ما يبدو متصل بالنواة المبهمة nucleus ambiguus ومن الممكن أن له تأثيرا مخفضا لضغط الدم depressor (بدليل ما ورد في هذه الدراسة مثلا).

التحديق الجانبي نحو اليسار يتطلب تنشيط مركز التحديق الجانبي على جهة اليسار. مركز التحديق الجانبي على جهة اليسار سينشط النواة القحفية رقم 6 على جهة اليسار والنواتين القحفيتين رقم 3 و4 على جهة اليمين.
الاتصال بين مركز التحديق الجانبي (الموجود في الجسر) وبين النواتين القحفيتين رقم 3 و4 (في الدماغ الأوسط) يتم عبر طريق يسمى “الحزمة الطولية الإنسية” (MLF) medial longitudinal fasciculus. لو تعرضت هذه الحزمة لأذى (مثلا بسبب مرض التصلب المتعدد multiple sclerosis أو بسبب سكتة دماغية stroke) فهذا يمكن أن يؤدي إلى خلل في التحديق الجانبي يسمى “الشلل العيني بين النووي” internuclear ophthalmoplegia. لو أصيبت الـ MLF على الجهة اليمنى فإن الإنسان سيكون عاجزا عن التحديق المزدوج نحو اليمين (العين اليمنى ستتحرك ولكن العين اليسرى لن تتحرك)، والعكس سيحصل لو أصيبت الـ MLF على الجهة اليسرى.

التحديق الإرادي

التحديق (كغيره من الحركات الهيكلية) يمكن أن يكون إراديا أو غير إرادي. التحديق الإرادي يتم بتأثير من ألياف نازلة من قشرة المخ نحو منظومة التحديق غير الإرادي الموجودة في جذع الدماغ.
المنطقة القشرية المسؤولة عن التحديق الإرادي تسمى “حقل العين الجبهي” (FEF) frontal eye field. موقع هذه المنطقة يختلف حسب المصادر. كثير من المصادر تقول أنها تقع في منطقةBrodmann رقم 8 في القشرة قبل الجبهية prefrontal cortex، ولكن هذه الدراسة تقول أنها تقع في منطقة Brodmann رقم 6 قرب القسم العلوي من الأخدود قبل المركزي precentral sulcus. هذا الوصف يعني أنها تقع في القشرة قبل المحركة premotor cortex.
كثير من المصادر تتحدث عن وجود مركز قشري خاص بحركة التعقب السلس smooth pursuit الإرادية اسمه “حقل التعقب الجبهي” frontal pursuit area، ولكن مكان هذا المركز هو غير واضح. حسب الدراسة التي أشرت إليها فإن “حقل التعقب الجبهي” هو القسم الخلفي من حقل العين الجبهي، وأما القسم الأمامي من حقل العين الجبهي فهو مسؤول عن حركة الطَّرْف saccade.
طبعا حركة التعقب السلس هي بطبيعتها حركة غير إرادية، ولكن التحفيز الكهربائي لـ “حقل التعقب الجبهي” ينشط هذه الحركة، ما يدل على وجود سيطرة إرادية عليها.
حسب بعض المصادر فإن التحديق العمودي الإرادي يتنشط قشريا بطريقة تختلف عن التحديق الأفقي الإرادي. التحديق العمودي الإرادي يتطلب على ما يبدو تنشيط مناطق أكبر في القشرة المخية مقارنة بالتحديق الأفقي الإرادي.
التحديق غير الإرادي
هناك لبس وعدم وضوح في آليات التحديق غير الإرادي في جذع الدماغ، ولكن ما يبدو لي هو أن هناك آليتين منفصلتين لكل من حركة الطَّرْف saccade وحركات العين الأخرى (التثبيت fixation والتعقب السلس smooth pursuit).
كل حركات العين غير الإرادية تخضع لتأثير التليلة العلوية superior colliculus. التليلة العلوية تنقسم إلى منطقتين، منطقة سطحية (ظهرية) ومنطقة عميقة (بطنية). المنطقة السطحية تتلقى أليافا من شبكية العين ومن القشرة البصرية في المخ، وعلى ما يبدو فإن خلايا هذه المنطقة تشكل خريطة ثنائية الأبعاد لمجال الرؤية على نحو شبيه بالخريطة الموجودة في شبكية العين. المنطقة العميقة من التليلة العلوية تحوي خريطة سمعية مصدرها ألياف قادمة من التليلة السفلية inferior colliculus، وتحوي خريطة حسية جسمانية somatosensory مصدرها ألياف قادمة من الحبل الشوكي تسمى “الطريق الشوكي-السقفي” spinotectal tract. كل هذه الخرائط الحسية تتكامل وتؤثر على “خريطة تحريكية” موجودة في المنطقة العميقة وظيفتها تحريك العينين في الفراغ. خلايا “الخريطة التحريكية” تتلقى أيضا أليافا من منطقة في قشرة المخ تقع بين الفصين القذاليoccipital والجداري parietal تسمى “حقل العين القذالي”occipital eye field أو “حقل العين الجداري” parietal eye field. هذه الألياف تنقل ربما المزيد من المعلومات الحسية الفراغية إلى خلايا الخريطة التحريكية.
الخلايا التحريكية ترسل أليافها نحو مراكز التحديق الأفقي والعمودي في الجسر والدماغ الأوسط. بعض الألياف تذهب أيضا نحو الخلايا المحركة في القسم الرقبي من الحبل الشوكي وتسمى “الطريق السقفي-الشوكي” tectospinal tract. الباحثون يعتقدون أن وظيفة الطريق السقفي-الشوكي هي إدارة الرأس تزامنا مع التحديق؛ لهذا السبب الباحثون يعرّفون التحديق gaze بأنه حركة مشتركة للعينين والرقبة. الطريق السقفي-الشوكي لدى بعض الفقاريات هو أكثر أهمية مما هو عليه لدى البشر: هذا الطريق يستطيع أن يحرك الجسم بكامله لدى الأسماك والطيور والفئران، ما يدل على أهمية سقف الدماغ الأوسط لدى تلك الكائنات.
التليلتان العلويتان ترسلان أيضا أليافا نحو الأنوية الجسرية العميقة deep pontine nuclei. هذه الألياف تسمى “الطريق السقفي-الجسري” tectopontine tract. الأنوية الجسرية العميقة ترسل أليافها نحو المخيخ. هناك اعتقاد بأن الطريق السقفي-الجسري يوصل معلومات الخريطة الحسية السقفية إلى المخيخ عبر الأنوية الجسرية العميقة.
هناك تشابه في تكوين ووظيفة سقف الدماغ الأوسط والمخيخ، وهذا ليس مستغربا نظرا لتشابه الأصل الجنيني لهذين التكوينين.
سقف الدماغ الأوسط (كبقية المراكز التحريكية في الدماغ الأوسط) يخضع لتأثير كابح من العقد القاعدية basal ganglia. هذا التأثير الكابح هو ضروري لتنظيم التحديق ومنعه من الانفلات. دون هذا الكبح فإن العينين سوف تحدقان باتجاه مؤثرات بصرية لا تستحق التحديق نحوها (ملهيات بصرية أو تشويش بصري).

التثبيت
الطرف saccade يعني تحريك العينين بأقصى سرعة ممكنة نحو نقطة معينة في مجال الرؤية. هذا التعريف لا يشمل تثبيت العينين على تلك النقطة fixation.
على ما يبدو فإن التثبيت غير الإرادي يتم تحت إشراف التليلة العلوية superior colliculus، بدليل وجود منطقة في التليلة العلوية مسؤولة عن التثبيت. هذه المنطقة تقع في القسم المنقاري من الخريطة التحريكية وهي تسمى “منطقة التثبيت” fixation zone أو “المنطقة النقرية”foveal zone (لأن تنشيطها يؤدي إلى تثبيت الصورة على النقرة fovea).
تنشيط منطقة التثبيت يثبط خلايا الخريطة التحريكية في التليلة العلوية، كما أنه ينشط نواة الرفاء الظهرية dorsal raphe التي تحوي خلايا مفرزة للـ GABA مسؤولة عن تثبيط حركات العين (تذكر ما قلناه سابقا عن الـ serotonin ودوره المثبط للحركة الهيكلية).
تثبيت الصورة على نقرة الشبكية fovea هو عملية صعبة لأن الرأس والجسم يتحركان باستمرار وهذا يؤدي لإزاحة الصورة عن النقرة. في جذع الدماغ هناك آلية خاصة وظيفتها تحريك العينين بما يحافظ على ثبات الصورة على النقرة. هذه الآلية تسمى المنعكس الدهليزي-العيني vestibulo-ocular reflex أو المنعكس العيني-الرأسي oculocephalic reflex.

تنسيق المنعكس الدهليزي-العيني يتم عبر النواة الدهليزية الإنسية medial vestibular nucleus ونواة قريبة منها تسمى nucleus prepositus hypoglossi. النواة الدهليزية الإنسية تتلقى دعما من ندفة المخيخ flocculus.

النواة الدهليزية الإنسية medial vestibular nucleus هي مسؤولة أيضا عن المنعكس الدهليزي-الرقبي vestibulocollic reflex الذي يحافظ على توازن الرأس عبر ألياف الطريق الدهليزي-الشوكي الإنسي medial vestibulospinal tract التي تذهب إلى الحبل الشوكي الرقبي (راجع هذا المقال للمزيد من المعلومات حول المنعكسات الدهليزية).
المنعكس الدهليزي-العيني هو ضروري لعملية التثبيت fixation التي تتم بإيعاز من التليلة العلوية. إتلاف النواة الدهليزية الإنسية لدى القرود لا يعيق حركة الطرف saccade ولكن القرد المصاب لا يستطيع أن يثبت عينيه على النقطة التي قام بالطرف نحوها (عيناه تطرفان نحو تلك النقطة ولكنهما سرعان ما تعودان إلى الوضع المركزي).
على ما يبدو فإن التليلة العلوية تثبت العينين عبر التأثير على المنعكس الدهليزي-العيني. لا أدري ما هي الآلية بالضبط، ولكن التليلة العلوية ترسل أليافا نحو أنوية جسرية “قبل مخيخية” precerebellar هي ما يلي:
- الأنوية الجسرية الوحشية الظهرية dorsolateral pontine nuclei (جزء من الأنوية الجسرية العميقة deep pontine nuclei)
- النواة الشبكية الغطائية الجسرية pontine reticulotegmental nucleus
الأنوية الجسرية الوحشية الظهرية ترسل أليافا نحو ندفة المخيخ flocculus، التي بدورها ترسل أليافا نحو الأنوية الدهليزية. هذا الطريق هو ربما صلة الوصل بين التليلة العلوية والمنعكس الدهليزي-العيني.
إذن تثبيت العينين غير الإرادي يمكن أن يبدأ من الجهاز الدهليزي أو من منطقة التثبيت في التليلة العلوية.
التعقب السلس
حركة التعقب السلس smooth pursuit (تسمى أيضا tracking) تشبه المنعكس الدهليزي-العيني. الفرق بين الحركتين هو أن الرأس في حركة التعقب السلس يكون ثابتا، وأما في المنعكس الدهليزي-العيني فالرأس يكون متحركا. في كلتا الحالتين العينان تتحركان بهدف تثبيت الصورة على النقرة fovea.
المنظومة الدماغية التي تنفذ حركة التعقب السلس هي ربما نفس المنظومة المسؤولة عن التثبيت والتي وصفت في الأعلى. الفرق بين التعقب السلس والتثبيت يتعلق ربما بالإشارات الواردة من الخلايا التحريكية في التليلة العلوية.
الرئيسيات (ومنها الإنسان) تستطيع أن تنفذ التعقب السلس الأفقي أو الجانبي بشكل أفضل من التعقب السلس العمودي. أيضا هي تستطيع أن تنفذ التعقب السلس نحو الأسفل بشكل أفضل من التعقب السلس نحو الأعلى. ربما يكون سبب هذا هو أن التعقب السلس الأفقي هو أهم تطوريا أو أقدم تطوريا. بعض الباحثين يرون أن التعقب السلس نشأ في الأصل بهدف مساعدة الحيوانات المفترسة على تعقب طرائدها.
خلاصة التحديق غير الإرادي
خلاصة ما سبق هي أن التحديق غير الإرادي يخضع لسيطرة التليلة العلوية. التليلة العلوية تنفذ حركة الطرف saccade عبر تأثيرها على مركز التحديق الجانبي في الجسر ومركز التحديق العمودي في الدماغ الأوسط المنقاري، وتنفذ حركتي التثبيت fixation والتعقب السلس smooth pursuit عبر تأثيرها على منظومة المنعكس الدهليزي-العيني vestibulo-ocular reflex.
من الأمور اللافتة أن إتلاف مركز التحديق الجانبي في الجسر ومركز التحديق العمودي في الدماغ الأوسط لا يلغي المنعكس الدهليزي-العيني ولا حركة التعقب السلس. هذا يدل على أن آلية المنعكس الدهليزي-العيني هي في الأصل مستقلة عن آلية الطرف saccade.
الاهتزازات المجهرية
تثبيت العينين الذي تحدثنا عنه في الأعلى هو أمر نسبي، لأن العينين في الحقيقة تهتزان مجهريا باستمرار، حتى عند التثبيت.
هناك اهتزاز مجهري مستمر لمقلة العين يسمى “الاهتزاز المجهري العيني” ocular microtremor. هذا الاهتزاز لا يتوقف إلا عند موت جذع الدماغ.
هناك نوع آخر من الاهتزاز العيني المجري يسمى “الطرف المجهري” microsaccade. الطرف المجهري هو عبارة عن حركات طرف مجهرية متتابعة تظهر عند تثبيت العينين. أول من لاحظ الطرف المجهري هو Robert Darwin والد العالم الشهير Charles Darwin.
بعض الباحثين يرون أن اهتزازات العين المجهرية تهدف لحماية المستقبلات الضوئية في الشبكية من التكيف adaptation.
التكيف هو ظاهرة عامة تصيب معظم المستقبلات الحسية. المقصود بالتكيف هو أن يتوقف المستقبل الحسي عن إرسال إشارات إلى الجهاز العصبي المركزي بعد مرور فترة معينة على بدء تنشيطه.
مثلا لو وضعنا يد إنسان في ماء بارد فإنه في البداية سيشعر بإحساس قوي بالبرودة وارد من يده، ولكن مع مرور الوقت سوف يخف إحساسه بالبرودة إلى أن يتلاشى تماما. هذه الظاهرة تحدث بسبب تكيف adaptation مستقبلات البرودة في جلد يده.
لو ثبتنا صورة معينة على المستقبلات الضوئية في الشبكية فهذا سيؤدي مع مرور الوقت إلى تكيف المستقبلات الضوئية، وبالتالي سوف تتوقف هذه المستقبلات عن إرسال الإشارات نحو الجهاز العصبي المركزي. لهذا السبب الاهتزازات المجهرية في العين هي ضرورية لمنع تكيف المستقبلات الضوئية.
في عام 1950 أجرى بعض الباحثين تجربة قاموا خلالها بتثبيت صورة معينة على الشبكية رغما عن الاهتزازات المجهرية. الشخص الذي خضع للتجربة شعر بأن الصورة التي يراها تلاشت تدريجيا ثم اختفت. هذا دليل على وجود التكيف في المستقبلات الضوئية في الشبكية.
ما أمام السقف
منطقة ما أمام السقف pretectum تقع في الجهة الظهرية من الدماغ الأوسط المنقاري على الجهة المنقارية للتليلتين العلويتين. أنا وصفت موقع هذه المنطقة في هذا المقال.
منطقة ما أمام السقف هي عبارة عن كتلتين تضمان عددا كبيرا من الأنوية. وظائف هذه الأنوية هي غير معروفة بشكل كامل، ولكنها تلعب دورا في تنسيق التحديق.
هناك ثلاثة منعكسات بصرية شهيرة يتم تنسيقها عبر ما أمام السقف. هذه المنعكسات هي:
- منعكس الضوء الحدقي pupillary light reflex
- منعكس التكيف accommodation reflex
- المنعكس البصري-الحركي optokinetic reflex
منعكس الضوء الحدقي

منعكس الضوء الحدقي pupillary light reflex هو ما يلي: عند سقوط ضوء قوي على شبكية العين فإن حدقتي العينين تتضيقان.
هذا المنعكس يتطلب تدخل نواةEdinger–Westphal المعروفة أيضا باسم “النواة المحركة العينية الملحقة” accessory oculomotor nucleus. أنا تحدثت عن العلاقة بين تضيق الحدقة pupil والضوء الساقط على العينين في هذا المقال.

منعكس التكيف

منعكس التكيف accommodation reflex يتنشط عند النظر إلى جسم قريب.
هذا المنعكس هو مركب من منعكسين:
- المنعكس الأول هو منعكس جانب متعاطف parasympathetic يتطلب تنشيط نواةEdinger–Westphal ويؤدي إلى تضيق الحدقتين وتحدب عدسة العين.
- المنعكس الثاني هو منعكس محرك جسماني somatic motor يتطلب تنشيط النواة فوق المحركة العينية supraoculomotor nucleus ويؤدي إلى مقاربة العينين convergence.
القشرة البصرية في المخ تلعب دورا في تنشيط هذا المنعكس، لأن هذا المنعكس يعتمد على تقدير البُعد.
أنا بينت سابقا العلاقة بين تضيق الحدقة وتحدب عدسة العين وبين رؤية الأجسام القريبة.
بالنسبة لمقاربة العينين convergence فهي ضرورية للمحافظة على جودة الرؤية ثنائية العين binocular vision، كما يظهر من الرسم التالي:

مقاربة العينين عند النظر إلى جسم قريب هي ضرورية لمنع الرؤية المزدوجة diplopia.
لو لم تتم مقاربة العينين عند النظر إلى جسم قريب فإن الصورة التي ستشاهدها كل من العينين ستكون مختلفة، وهذا سيؤدي للرؤية المزدوجة.
الرؤية المزدوجة تنشأ لدى كل إنسان عندما يقرب إصبعه من أنفه وهو ينظر إليه بكلتا عينيه. عندما يقترب الإصبع كثيرا من الأنف فسوف يصبح إصبعين وليس إصبعا واحدا، لأن كل عين سترى الإصبع بصورة مختلفة.
بعض الباحثين يرون أن سبب تسطح الوجه عند الرئيسيات primates هو زيادة قدرتها على مقاربة العينين.

حركة المقاربة convergence تتطلب تحريك العينين نحو الداخل، أي باتجاه الأنف. لو كان الوجه والأنف بارزين نحو الأمام فهذا سيقلل من مجال الرؤية ثنائية العين للأجسام القريبة.
الرئيسيات الباكرة كانت على ما يبدو تحتاج للرؤية ثنائية العين على مسافة قريبة، ولهذا السبب تسطحت وجوهها لإفساح المجال أمام قدر كبير من المقاربة convergence.

المنعكس البصري-الحركي
المنعكس البصري-الحركي optokinetic reflex هو منعكس مركب من الطرف saccade والتعقب السلس smooth pursuit.
لو كنت راكبا في سيارة وتراقب الأشجار على جانب الطريق فإن عينيك ستنفذان المنعكس البصري-الحركي. في البداية عيناك ستنفذان حركة طرف باتجاه شجرة معينة وستتثبتان عليها. بما أن السيارة تسير فإن الشجرة ستتحرك تدريجيا إلى خارج مجال الرؤية. عيناك ستلاحقان الشجرة بالتعقب السلس إلى أن تخرج من مجال الرؤية، وبعد ذلك ستعود عيناك بواسطة حركة طرف إلى نفس الوضعية التي كانتا عليها عند بداية رصد الشجرة.
هذا المنعكس يمكن أن يتنشط دونما سبب ظاهر، وعندها فإنه يسمى nystagmus.
الدماغ البشري (14)

ذكرنا سابقا أن جميع الأنوية القحفية الموجودة في الدماغ الأوسط هي مسؤولة عن تحريك العينين باستثناء نواة الدماغ الأوسط للعصب القحفي الخامس mesencephalic nucleus of trigeminal nerve، التي هي نواة حسية جسمانية somatic sensory تستقبل الحس الذاتي (غير الواعي؟) من النصف الأمامي لجدار الرأس (الوجه والجبهة والذقن).

في مقطع الدماغ الأوسط الذيلي caudal midbrain تظهر نواة العصب القحفي الرابع أو البكري trochlear nerve. هذه النواة تؤدي وظيفة مكملة لعمل نواة العصب القحفي الثالث (المحرك العيني oculomotor nerve) الموجودة في الدماغ الأوسط المنقاري rostral midbrain. العصب القحفي الرابع يحرك عضلة واحدة فقط من العضلات المحركة لمقلة العين هي العضلة المائلة العلوية superior oblique muscle. وظيفة هذه العضلة هي تحريك مقلة العين في الاتجاه الظاهر في هذه الصورة:

إذا أصيب أحد العصبين البكريين بأذى فسوف تكون هناك صعوبة في تحريك إحدى العينين نحو الأسفل والداخل، وهذا سيؤدي إلى الرؤية المزدوجة diplopia عندما ينظر المصاب في اتجاه يستدعي تحريك عينه المصابة نحو الأسفل أو الداخل.


في مقطع الدماغ الأوسط المنقاري rostral midbrain تظهر أنوية العصب القحفي الثالث (العصب المحرك العيني oculomotor nerve). هذا العصب هو مسؤول عن تحريك جميع عضلات العين ما عدا عضلتين:
- العضلة المائلة العلوية superior oblique muscle (يحركها العصب القحفي الرابع أو البكري trochlear)
- العضلة المستقيمة الوحشية lateral rectus muscle (يحركها العصب القحفي السادس أو المبعد abducens)

نواة العصب القحفي الثالث هي ليست نواة واحدة وإنما مجموعة من الأنوية (تسمى أحيانا “المجمّع المحرك العيني” oculomotor complex). هذه الأنوية هي جسمانية محركة somatic motor وظيفتها تحريك العضلات المحركة للعين. على الجهة الظهرية للمجمع المحرك العيني هناك نواة حشوية محركة visceral motor تسمى “النواة المحركة العينية الملحقة” accessory oculomotor nucleus أو Edinger–Westphal nucleus. هذه النواة هي جزء من الجهاز العصبي جانب المتعاطف parasympathetic وهي مصدر الألياف الحشوية جانب المتعاطفة التي تصل إلى العين عبر العصب المحرك العيني والعقدة الهدبية ciliary ganglion. هناك أيضا ألياف متعاطفة sympathetic تصل إلى العين من العقدة الرقبية العلوية superior cervical ganglion (التي تتلقى أليافا قبل عقدية preganglionic مصدرها القسم الصدري من الحبل الشوكي).

التعصيب الحشوي أو المستقل autonomic يؤثر على جزأين من العين هما الحدقة pupil والعدسة lens.

دور الحدقة في الرؤية
التعصيب المتعاطف sympathetic يوسع الحدقة، والتعصيب جانب المتعاطف parasympathetic يضيقها. توسيع الحدقة هو مناسب لرؤية الأجسام البعيدة، وتضييق الحدقة هو مناسب لرؤية الأجسام القريبة.

الجسم القريب يرسل أشعة ضوئية نحو مساحة واسعة من شبكية العين retina، بخلاف الجسم البعيد. هذا الأمر يضعف من حدة الرؤية، لأن المناطق الطرفية من الشبكية تتميز بقلة الدقة مقارنة بالمناطق المركزية.
هناك نوعان من المستقبلات الضوئية في شبكية العين: العصي rods والمخاريط cones. العصي هي قليلة الدقة مقارنة بالمخاريط، ولكنها تعمل بشكل أفضل من المخاريط في الضوء الخافت. المخاريط هي أكثر دقة من العصي، ولكنها لا تعمل بشكل جيد في الضوء الخافت.

المخاريط تتركز في وسط الشبكية، وخاصة في النقطة المسماة “النُّقْرة المركزية” fovea centralis. هذه النقطة تخلو من العصي وتحوي كما هائلا من المخاريط. النقرة هي مصدر 50% من ألياف العصب البصري optic nerve الذي ينقل حس البصر إلى الدماغ.
نسبة المخاريط تقل تدريجيا كلما ابتعدنا عن النقرة المركزية. المناطق الطرفية من الشبكية تحوي عددا قليلا من المخاريط، ولذلك هي قليلة الدقة مقارنة بالمنطقة المركزية.
العين في الأحوال الطبيعية تحاول أن توجه الضوء الساقط على العين نحو النقرة المركزية، لأن النقرة المركزية (والمنطقة المحيطة بها) هي أفضل مناطق الشبكية للرؤية الدقيقة أو الحادة.
أشعة الضوء القادمة من جسم بعيد عن العين تسقط على القسم المركزي من الشبكية، ولكن تقريب الجسم من العين سيؤدي إلى سقوط أشعة الضوء على المناطق المحيطية من الشبكية (انظر الرسم في الأعلى). سقوط الضوء على المناطق المحيطية من الشبكية سينشط الكثير من العصي ذات الدقة الضعيفة، وهذا سيشوش الرؤية وسيجعلها غير حادة أو “مغبشة” blurred.

تضييق حدقة العين عند النظر إلى الأجسام القريبة يخفف من “التغبيش” ويزيد من حدة الرؤية، لأنه يحصر سقوط الأشعة الضوئية على المنطقة المركزية من الشبكية حيث تكثر المخاريط.
هذا الكلام هو صحيح فقط عندما تكون هناك إضاءة جيدة، لأن المخاريط لا تعمل دون إضاءة جيدة. إذا كانت الإضاءة خافتة فإن توسيع الحدقة عند النظر إلى الأجسام القريبة يصبح أمرا جيدا، لأنه يزيد من كمية المستقبلات الضوئية الناشطة (التي هي العصي في هذه الحالة).
المخاريط تملك القدرة على التمييز بين الألوان، ولكن العصي لا تملك هذه القدرة. لهذا السبب الإنسان يستطيع أن يميز الألوان فقط عندما تكون هناك إضاءة جيدة. عندما تخفت الإضاءة كثيرا فإن الإنسان يرى كل شيء باللون الرمادي.
التعصيب المتعاطف sympathetic يوسع الحدقة، ولذلك هو مفيد لرؤية الأجسام البعيدة والرؤية في الظلام. التعصيب جانب المتعاطف parasympathetic يضيق الحدقة، ولذلك هو مفيد لرؤية الأجسام القريبة والرؤية في الإضاءة الجيدة.
دور العدسة في الرؤية
التعصيب المتعاطف يقلل من تحدب العدسة، والتعصيب جانب المتعاطف يزيد من تحدب العدسة. تقليل تحدب العدسة هو مناسب لرؤية الأجسام البعيدة، وزيادة تحدب العدسة هي مناسبة لرؤية الأجسام القريبة.
عدسة العين هي عدسة محدبة convex. بؤرة العدسة المحدبة هي النقطة التي تجتمع عندها أشعة الضوء بعد المرور بالعدسة.

بُعد البؤرة focus عن العدسة يعتمد على عاملين: شدة تحدب العدسة، واتجاه الأشعة الضوئية الساقطة على العدسة.
الأجسام القريبة من العدسة ترسل أشعة ضوئية متباعدة الاتجاهات. هذا يؤدي إلى زيادة بعد البؤرة عن العدسة. الأجسام البعيدة من العدسة ترسل أشعة ضوئية متوازية. هذا يؤدي إلى تقريب البؤرة من العدسة.

عدسة العين تحاول أن توجه الضوء الساقط عليها نحو النقرة fovea. لهذا السبب هي تحاول أن تجعل بؤرتها في النقرة. التعصيب المستقل (المتعاطف وغير المتعاطف) يغير من شدة تحدب العدسة بهدف جعل بؤرتها في النقرة. هذه العملية تسمى “التكيف” accommodation. عند النظر إلى جسم قريب close object يقوم التعصيب جانب المتعاطف بزيادة تحدب العدسة لكي يمنع ابتعاد البؤرة إلى ما خلف الشبكية. عند النظر إلى جسم بعيد distant object يقوم التعصيب المتعاطف بتقليل تحدب العدسة لكي يمنع اقتراب البؤرة إلى ما أمام الشبكية.
الخلل في قدرة العين على التكيف accommodation هو سبب “قصر النظر” myopia و”بعد النظر” hyperopia.

تكيف عدسة العين accommodation هو نفس مفهوم “الفوكسة” to focus. الخلل في “فوكسة” عدسة الكاميرا يجعل الصورة “مغبشة” blurred. هذه هي نفس المشكلة التي تحصل لدى المصابين بقصر النظر أو بعد النظر.
الرؤية ثنائية العين
الفقاريات عموما تملك عينين اثنتين. كل عين من العينين لها حقل رؤية visual field. هناك في الغالب تداخل بين حقلي الرؤية للعينين. منطقة التداخل بين حقلي الرؤية تسمى حقل “الرؤية ثنائية العين” binocular vision.

التداخل بين حقلي الرؤية للعينين يفيد في إدراك البُعد depth perception أو الرؤية المجسمة stereopsis. إدراك البعد يفيد الحيوانات المفترسة predatory، ولهذا السبب الرؤية ثنائية العين تكثر لدى الحيوانات المفترسة. بالنسبة للفرائس preys فإن امتلاك مجال رؤية واسع هو أهم من القدرة على إدراك البعد، ولهذا السبب الرؤية ثنائية العين هي عموما قليلة لدى مثل هذه الحيوانات.

الرئيسيات primates تتميز بامتلاكها مجالا واسعا للرؤية ثنائية العين، رغم أن كثيرا من الرئيسيات هي ليست حيوانات مفترسة، والرئيسيات الباكرة لم تكن على الأغلب حيوانات مفترسة.

بعض الباحثين يعتقدون أن الرئيسيات الباكرة كانت تعيش على أغصان الأشجار، والرؤية ثنائية العين هي مفيدة للحياة في هكذا بيئة، لأن التقدير الصحيح للبعد يسهل القفز من غصن إلى آخر ويمنع السقوط على الأرض (ولكن هذه النظرية هي مثيرة للجدل. للمزيد انظر هذا المقال).
الرؤية ثنائية العين تستلزم توجيه العينين نحو نفس الهدف، وإلا فإن المرء سيعاني من الرؤية المزدوجة diplopia ومن ضعف في إدراك البعد (كما يحصل مثلا عند الإصابة بالحول strabismus).
الدماغ البشري (13)
تحدثت في السابق عن “ساقي المخ” crura cerebri اللتين تقعان على الجهة البطنية للدماغ الأوسط (midbrain) mesencephalon والدماغ البيني diencephalon (واللتين تتماديان مع “المحفظتين الداخليتين” internal capsules في المخ cerebrum)، وتحدثت عن منطقة “الغطاء” tegmentum التي تقع على الجهة الظهرية dorsal لساقي المخ (والتي تحوي التشكل الشبكي reticular formation).
على الجهة الظهرية للغطاء توجد الأنوية القحفية cranial nuclei. جميع الأنوية القحفية الموجودة في الدماغ الأوسط هي مسؤولة عن تحريك العينين باستثناء نواة الدماغ الأوسط للعصب القحفي الخامس mesencephalic nucleus of trigeminal nerve. هذه النواة هي نواة حسية جسمانية somatic sensory تستقبل الحس الذاتي proprioception (غير الواعي؟) القادم من النصف الأمامي لجدار الرأس (الوجه والجبهة والذقن) عبر العصب القحفي الخامس (العصب ثلاثي التوائم trigeminal nerve). الأصل الجنيني لهذه النواة يعود إلى الجسر pons في الدماغ الخلفي (hindbrain) rhombencephalon.
في مقطع الدماغ الأوسط الذيلي تظهر نواة قحفية أخرى هي نواة العصب القحفي الرابع (العصب البكري trochlear nerve). هذه النواة هي نواة محركة جسمانية somatic motor تساهم في تحريك مقلة العين. الأصل الجنيني لهذه النواة يعود أيضا إلى الجسر.

في مركز الدماغ الأوسط توجد “قناة الماء الدماغية” (“المَسال الدماغي”) cerebral aqueduct. هذه القناة هي استمرار للبطين الرابع fourth ventricle الموجود في الدماغ الخلفي.

أنا وضعت سابقا هذه الصورة التي تبين كيفية اتساع القناة المركزية في الحبل الشوكي وتحولها إلى البطين الرابع:

آلية تحول البطين الرابع fourth ventricle إلى قناة الماء الدماغية cerebral aqueduct هي معاكسة للآلية الظاهرة في الصورة أعلاه.

قناة الماء الدماغية تتسع مجددا في الدماغ البيني diencephalon وتتحول إلى البطين الثالثthird ventricle . البطين الثالث يبدو كبيرا عندما ننظر له من الجهة الجانبية أو الوحشية lateral، ولكن لو نظرنا إليه من الأمام أو الخلف فسيتبين أنه ضيق.
آلية تحول قناة الماء الدماغية إلى البطين الثالث تشبه نوعا ما آلية تحول قناة الحبل الشوكي المركزية إلى البطين الرابع. ما يحصل هو أن قناة الماء تنفتح من الجهة الظهرية dorsal، ولكن في الدماغ البيني لا يوجد دوران للمادة الرمادية نحو الجهة الوحشية lateral.
ما يلي مقطع عرضي في الدماغ البيني. تذكر أن الدماغ البيني محاط بالمخ، وبالتالي من غير الممكن أخذ مقطع عرضي في الدماغ البيني لا يظهر فيه المخ أيضا:

ما يلي صورة مقربة:

لاحظ أن البطين الثالث في المقطع العرضي يأخذ شكل شق طولي، ولاحظ أن سقف البطين الثالث مكون من أجزاء تنتمي إلى المخ cerebrum (الدماغ النهائي telencephalon).
إذن البطين الثالث هو محاط من جميع الجهات بأجزاء تنتمي إلى الدماغ البيني ما عدا الجهة العلوية (التي هي أصلا الجهة الظهرية للأنبوب العصبي).
الجهة العلوية للدماغ البيني هي أصلا مفتوحة (مغطاة فقط بغشاء ظهاري رقيق)، ولكن المخ التف نحو الخلف وغطاها. تذكر أن المخ هو ملتف نحو الخلف كما يظهر من الرسم التالي:

لنعد مجددا إلى قناة الماء الدماغية cerebral aqueduct. حول قناة الماء الدماغية توجد كتلة من المادة الرمادية تسمى “المادة الرمادية حول قناة الماء” (PAG) periaqueductal gray (تسمى أيضا “المادة الرمادية المركزية” central gray). من الناحية الجنينية هذه المادة هي مشتقة من الصفيحتين الجناحيتين alar plates (أي من القسم الظهري للأنبوب العصبي)، ومن الناحية الوظيفية هي تعتبر امتدادا ذيليا لـ “ما تحت الثلم” hypothalamus (الذي هو أيضا مشتق من الصفيحتين الجناحيتين).

على الجهة الظهرية للـ PAG يوجد السقف tectum.
لو نظرنا إلى السقف من الجهة الظهرية فسنجد أنه يتكون من أربعة نتوءات.


كل نتوء من نتوءات السقف الأربعة يسمى “تُلَيْلة” colliculus (كلمة colliculus هي تصغير لـcollis التي تعني “تلة” hill). هناك تليلتان علويتان superior colliculi وتليلتان سفليتان inferior colliculi.
التليلات الأربع تسمى أحيانا “الأجسام التوأمية الأربعة” corpora quadrigemina.
التليلتان العلويتان تستقبلان أليافا تنقل حس البصر قادمة من الطريق البصري optic tract عبر طريق يسمى “ذراع التليلة العلوية” brachium of the superior colliculus. هذا الطريق يربط التليلتين العلويتين مع “الجسم الركبي الوحشي” lateral geniculate body الموجود في مؤخرة الثلم thalamus.

التليلتان السفليتان تستقبلان أليافا تنقل حس السمع قادمة من النواتين القوقعيتين cochlear nuclei عبر “الشريط الوحشي” lateral lemniscus. التليلتان السفليتان تصدران أليافا نحو “الجسم الركبي الإنسي” medial geniculate body عبر طريق يسمى “ذراع التليلة السفلية” brachium of the inferior colliculus.

على الجهة المنقارية rostral للتليلتين العلويتين تقع المنطقة المسماة “ما أمام السقف” pretectum أو “المنطقة أمام السقفية” pretectal area. هذه المنطقة هي مكونة من كتلتين متناظرتين من الأنوية على اليمين واليسار. لو أخذنا مقطعا عرضيا في القسم المنقاري الأقصى من الدماغ الأوسط فسنجد كتلتي “المنطقة أمام السقفية” في مكان التليلتين العلويتين. المنطقة أمام السقفية هي مسؤولة عن تنسيق منعكسات بصرية. على الجهة البطنية ventral لهذه المنطقة يوجد القسم المنقاري الأقصى من الـ PAG.
على الجهة المنقارية للمنطقة أمام السقفية يوجد “الملتقى الخلفي” posterior commissure، الذي هو شريط من الألياف العصبية المعترضة التي تسير من اليمين إلى اليسار ومن اليسار إلى اليمين. على الجهة المنقارية للملتقى الخلفي توجد الغدة الصنوبرية pineal gland. الغدة الصنوبرية لها ساقان يمنى ويسرى. كل ساق من هاتين الساقين تسمى “العُنَيِّن” habenula (كلمة habēnula اللاتينية هي تصغير لـ habēna التي تعني “العِنَان” أو “الزِّمَام” أو “اللِّجام” أو “الرَّسَن”، أي الحبل الذي تربط به الدابة. كلمة “عُنَيِّن” هي تصغير لـ “عِنان”).
العنين هو شريط من الألياف العصبية، ولكنه يحوي في قاعدته كتلة من المادة الرمادية تسمى “النواة العنينية” habenular nucleus. عندما ترد كلمة habenula في الكتابات المعاصرة فإن المقصود بها على الأغلب هو النواة العنينية، وأما الألياف التي تذهب نحو الغدة الصنوبرية فهي تسمى “ساق الغدة الصنوبرية” stalk of pineal gland.
هناك اتصال بين منطقة النواة العنينية اليمنى ومنطقة النواة العنينية اليسرى على الجهة المنقارية لساقي الغدة الصنوبرية. هذا الاتصال يسمى “الملتقى العنيني” habenular commissure.

القسم المنقاري الأقصى من غطاء الدماغ الأوسط midbrain tegmentum يشكل جزءا من أرضية البطين الثالث ومن جدرانه الجانبية السفلية، كما يظهر في الرسم التالي:

لو أخذنا مقطعا عرضيا في القسم الخلفي من الدماغ البيني فسنرى في أسفل المقطع أجزاء غطاء الدماغ الأوسط المنقاري (من قبيل النواة الحمراء red nucleus والمادة السوداء substantia nigra). السبب ظاهر في الرسم أعلاه.
العنين habenula يتصل في الجهة الأمامية مع شريط من الألياف العصبية يسمى “الخط النخاعي للثلم” stria medullaris thalami. هذا الخط يسير على الجانب الإنسي medial للثلم بمحاذاة “شريط الثلم” taenia thalami.
الرسم التالي يبين موقع “شريط الثلم” taenia thalami.
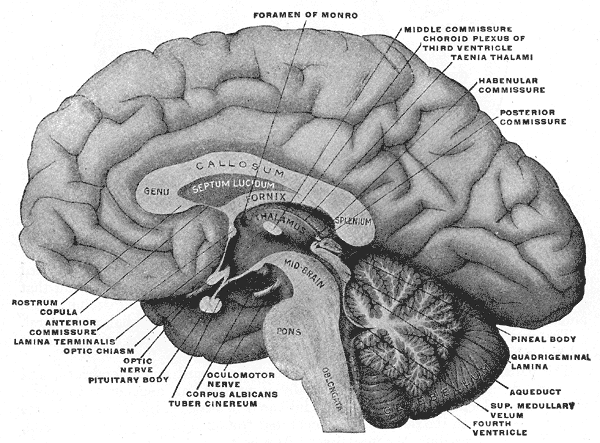
شريط الثلم taenia thalami يمثل الحد العلوي للجدار الجانبي للبطين الثالث. سطح الثلم الذي يقع فوق مستوى شريط الثلم هو جزء من أرضية البطين الجانبي lateral ventricle (الذي يقع في قلب نصف الكرة المخية cerebral hemisphere).
ما يلي مقطع عرضي في البطين الثالث والبطينين الجانبيين. لاحظ أن سطح الثلم thalamus يمثل جزءا من الجدار الجانبي للبطين الثالث وجزءا من أرضية البطين الجانبي، ولاحظ أن الضفائر المشيمائية choroid plexuses تقع في سقف البطين الجانبي وفي أرضية البطينين الجانبيين.

ما يلي رسم آخر يبين موقع العنين والشريط النخاعي stria medullaris:

ما يلي منظر ظهري:
الغدة الصنوبرية pineal gland والعنين habenula والملتقي العنيني habenular commissure والخط النخاعي stria medullaris تسمى جميعا “ما فوق الثلم” epithalamus. “ما فوق الثلم” هو جزء من الدماغ البيني diencephalon.
الملتقى الخلفي posterior commissure والمنطقة أمام السقفية pretectal area تعتبر من أجزاء الدماغ الأوسط.
الدماغ البشري (12)
في المقال الأخير من هذه السلسلة تحدثت عن أنواع الحركة الانتقالية locomotion لدى الحيوانات، وبينت أن نوع الحركة الانتقالية الذي كان سائدا لدى الفقاريات البدائية هو الحركة التموجية undulatory locomotion. عندما ظهرت الفقاريات رباعية الأرجل tetrapods ظهر نوع جديد من الحركة الانتقالية يعتمد على الأطراف المُمَفْصلة jointed limbs. الحركة الانتقالية بواسطة الأطراف الممفصلة تشمل مثلا حركات المشي والركض والقفز والخفق بالأجنحة.
تنفيذ الحركة الانتقالية التموجية يعتمد على العضلات الباسطة المحوريةaxial extensor muscles التي يؤدي تقليصها إلى بسط الجسم. في مقالات سابقة بينت أن هناك أنوية في التشكل الشبكي reticular formation في الدماغ الخلفي (hindbrain) rhombencephalon يؤدي تنشيطها إلى تقليص أو إرخاء العضلات الباسطة المحورية. هذه الأنوية هي ما يلي:
- النواة الشبكية عملاقة الخلايا gigantocellular reticular nucleus (في النخاع المستطيل)
- النواة الشبكية الجسرية الذيلية caudal pontine reticular nucleus (في الجسر الذيلي)
- النواة الشبكية الجسرية الفموية oral pontine reticular nucleus (في الجسر المنقاري)
هذه الأنوية ترسل أليافا نازلة نحو القرنين البطنيين للحبل الشوكي عبر الطريق الشبكي-الشوكي reticulospinal tract. من خلال هذه الألياف يمكن لهذه الأنوية أن تثير أو تثبط الخلايا المحركة الجسمانية somatic motor المسؤولة عن تقليص العضلات الباسطة المحورية.
في المقال الأخير تحدثت عن النواة الحمراء كبيرة الخلايا magnocellular red nucleus الموجودة في القسم المنقاري من الدماغ الأوسط rostral midbrain. هذه النواة ترسل أليافا نازلة نحو القرنين البطنيين للحبل الشوكي عبر الطريق الأحمر-الشوكي rubrospinal tract. هذه الألياف تصل لدى القطط (وغيرها من الثدييات) نحو القسم القَطَني lumbar من الحبل الشوكي حيث توجد الخلايا المحركة الجسمانية المسؤولة عن تقليص عضلات الطرفين الخلفيين (الرجلين)، ولكنها لدى البشر لا تنزل أبعد من القسم الرَقَبيcervical من الحبل الشوكي حيث توجد الخلايا المحركة الجسمانية المسؤولة عن تقليص عضلات الطرفين الأماميين أو العلويين (الذراعين). بعض الباحثين يعتقدون أن النواة الحمراء كبيرة الخلايا هي مسؤولة عن الحركة الانتقالية بواسطة الأطراف الممفصلة. الضمور الذي أصاب هذه النواة وأليافَها لدى البشر ربما يعود إلى تغير طريقة الحركة الانتقالية بواسطة الأطراف الممفصلة لدى البشر: البشر يعتمدون على المشية ثنائية القدم bipedal وليس المشية رباعية القدمquadrupedal كبقية الفقاريات رباعية الأرجل (للمزيد من المعلومات حول النواة الحمراء راجع المقال السابق).

إيقاع الحركة الانتقالية
أحد العناصر المهمة للحركة الانتقالية لدى الفقاريات هو الإيقاع rhythm. من يراقب الحركة الانتقالية التموجية أو الحركة الانتقالية بواسطة الأطراف الممفصلة سوف يلاحظ أن هذه الحركات هي حركات إيقاعية rhythmic، بمعنى أنها تتكون من حركات تتكرر باستمرار.
الحركة التموجية هي عبارة عن تكرار لحركة شبيهة بالموجة الجيبية sine wave (ومن هنا جاءت تسميتها بالحركة التموجية).

تردد الموجة الجيبية frequency يساوي حاصل قسمة السرعة التي تنتقل بها الموجة (سرعة الطور phase velocity) على طول الموجة wavelength.
حركة المشي هي عبارة عن تكرار لحركة تسمى الخطوة step. الباحثون يقسمون “حلقة الخطو” step cycle إلى طورين:
- طور الوقوف stance phase أو طور البسط extension phase (عندما تكون القدم ملامسة للأرض)
- طور التأرجح swing phase أو طور الثني flexion phase (عندما تكون القدم غير ملامسة للأرض)
هذه الصورة المتحركة تبين حركة المشي لدى الفيل:

الباحثون يقيسون حركة المشي في العادة بناء على حركة الطرف الخلفي الأيسر. عندما يكمل الطرف الخلفي الأيسر حلقة الخطو (طور الوقوف + طور التأرجح) فإن هذه تعتبر مشية كاملة. إذن لو أردنا أن نقيس تردد المشي لدى كائن ما فيجب علينا أن نقيس الزمن الذي تستغرقه حلقة الخطو للطرف الخلفي الأيسر.
الباحثون لاحظوا أن تردد أو سرعة المشي يعتمد في الأساس على الفترة الزمنية التي يستغرقها طور الوقوف. طور التأرجح هو عموما ثابت ولا يتغير كثيرا سواء كانت المشية سريعة أم بطيئة. ما يتغير في الأساس هو طور الوقوف. المشي السريع أو الركض يتم عبر تقليص الفترة الزمنية التي يستغرقها طور الوقوف.
الحركة الانتقالية الشوكية
الباحثون لاحظوا منذ القرن 19 أن الحبل الشوكي هو قادر بمفرده على إنتاج حركة المشي، دون تدخل من الدماغ.
هذا الاكتشاف ظهر من التجارب على الحيوانات “المُشَوَّكة” spinalized. المقصود بكلمة “التشويك” spinalization هو فصل الدماغ عن الحبل الشوكي. الحيوان الذي تم قطع دماغه بالكامل بحيث لم يبق من جهازه العصبي المركزي سوى الحبل الشوكي يسمى حيوانا مُشَوَّكًا spinalized.
الباحثون لاحظوا أن الحيوانات المشوكة تستطيع أن تمشي. هم حفزوا الأطراف الخلفية لحيوانات مشوكة وهذا أدى لتحركها على نحو شبيه بالمشي. لهذا السبب الباحثون رأوا أن حركة المشي هي منعكس شوكي.
الكلام التالي مأخوذ من كتاب نشر في عام 1879 للمؤلف Michael Foster:
. . . the [spinal] cord contains a number of more or less complicated mechanisms capable of producing, as reflex results, co-ordinated movements(?) altogether similar to those which are called forth by the will. Now it must be an economy to the body, that the will should make use of these mechanisms already present, by acting directly on their centers, rather than it should have recourse to a special apparatus of its own of a similar kind.
Cited in Wilfrid Jänig (2006), The Integrative Action of the Autonomic Nervous System
في القرن العشرين تبين أن الحركة التموجية للفقاريات البدائية (مثلا سمكة اللامبري lamprey) يمكن أيضا أن تنتج من الحبل الشوكي دون تدخل الدماغ. التجارب على سمكة اللامبري “المُشَوَّكة” أثبتت ذلك.
إذن حركتا التموج والمشي هما مزروعتان في الحبل الشوكي على نحو شبيه بالمنعكسات الشوكية، ولكن الباحثين حاليا (حسب علمي) لا يصنفون هاتين الحركتين ضمن المنعكسات الشوكية. السبب الأساسي لذلك هو أن هاتين الحركتين هما حركتان إيقاعيتان بخلاف المنعكسات الشوكية البسيطة.

المنعكس الشوكي البسيط هو عبارة عن دارة circuit. لكي يحدث رد الفعل لا بد من أن يكون هناك فعل. الفعل الواحد ينتج رد فعل واحدا. لكي يتكرر رد الفعل لا بد من تكرار الفعل.
منعكسا التموج والمشي لا يعملان بهذه الطريقة. لو قمنا بتنشيط هذين المنعكسين فإنهما سيظلان ناشطين بعد انقطاع المؤثر الحسي الذي أدى لتنشيطهما. هذا يدل على وجود إيقاع ذاتي أو أوتوماتيكي لهذين المنعكسين.
الباحثون حاليا يعتقدون أن هناك شبكات عصبية في الحبل الشوكي (مكونة من خلايا ربطية interneurons) تقوم بتوليد إيقاع التموج والمشي. هذه الشبكات لا تحتاج لمؤثر حسي لكي تستمر في العمل، أي أنها تولد الإيقاع بشكل ذاتي أو أوتوماتيكي.
بعض المؤلفين (Wilfrid Jänig) أطلقوا على هذه الشبكات مسمى “البرامج المحرّكة الشبكية” spinal motor programs، ولكن التسمية التي ترد في غالبية المصادر هي “مولدات النمط المركزية” central pattern generators.
المقصود بمولد النمط المركزي هو شبكة عصبية قادرة على توليد إيقاع تلقائي دون الحاجة لمؤثر حسي. أهم مولدات النمط المركزية لدى الفقاريات هي ما يلي:
- مولدات نمط الحركة الانتقالية locomotion pattern generators (في الحبل الشوكي)
- مولدات نمط التنفس respiration pattern generators (في النخاع المستطيل)
- مولدات نمط البلع swallowing pattern generators (في النخاع المستطيل)
مولدات نمط الحركة الانتقالية هي ممتدة على طول الحبل الشوكي لدى سمكة اللامبري، ولكنها لدى الحيوانات رباعية الأرجل تتركز في المنطقتين الرقبية cervical والقطنية lumbar. المنطقة الرقبية من الحبل الشوكي تحوي الخلايا المحركة المسؤولة عن قبض وبسط الطرفين الأماميين، والمنطقة القطنية تحوي الخلايا المحركة المسؤولة عن قبض وبسط الطرفين الخلفيين.
مولدات نمط الحركة الانتقالية الموجودة في المنطقة القطنية هي التي تحدد إيقاع المشي لدى الحيوانات ذات المشية رباعية القدمquadruped . هناك ألياف صادرة من مولدات النمط الموجودة في المنطقة القطنية نحو مولدات النمط الموجودة في المنطقة الرقبية، ولكن ليس العكس. هذا يعني أن مولدات النمط الرقبية تتبع في إيقاعها مولدات النمط القطنية.
الخلايا المكونة لمولدات نمط الحركة الانتقالية هي خلايا ربطية interneurons من نوعين:
- خلايا إثارية excitatory مفرزة للـ glutamate
- خلايا تثبيطية inhibitory مفرزة للـ glycine
هذه الخلايا تتلقى أليافا واردة من مصدرين:
- ألياف محيطية واردة من المستقبلات الحسية في الجلد والعضلات الهيكلية والمفاصل.
- ألياف مركزية واردة من الدماغ (عبر الطريق القشري-الشوكي corticospinal والطريق الشبكي-الشوكيreticulospinal والطريق الدهليزي-الشوكيvestibulospinal والطريق الأحمر-الشوكي rubrospinal).
الألياف المحيطية الحسية تلعب دورا هاما في تنظيم عمل مولدات نمط المشي. مثلا لو مشت الأقدام على أرض مغطاة بالحصى فهذا سينشط المستقبلات الحسية الموجودة في الجلد، والإشارات الواردة من هذه المستقبلات ستؤدي إلى تعديل المشية. أيضا إشارات الحس الذاتي proprioception الواردة من المستقبلات الحسية في العضلات الهيكلية والمفاصل تؤدي إلى تعديل سرعة وشدة تقلص عضلات الأطراف بهدف المحافظة على ثبات واتزان المشية.
المشي هو في الحقيقة منعكس مركب من مجموعة كبيرة من المنعكسات المتداخلة والمتراكبة (على غرار منعكسي التنفس والبلع وغيرهما من المنعكسات المركبة). من الأمور اللافتة أن خلايا مولدات نمط المشي تؤدي وظائف أخرى إلى جانب وظيفة توليد نمط المشي:
[The] spinal motor programs [=central pattern generators] are not static in the sense that one population of interneuron is involved in only one motor program (e.g., spinal locomotion), but rather is dynamic. Thus, individual groups of interneurons may be part of several spinal motor programs depending on the synaptic afferent inputs and on the behavioral state of the organism.
Wilfrid Jänig (2006), The Integrative Action of the Autonomic Nervous System p. 364
المقصود بهذا الكلام هو أن مجموعات معينة من الخلايا المولدة لنمط المشي يمكن أن تساهم في توليد أنماط أخرى من الحركة. هذا يدل على أن “نمط المشي” هو في الحقيقة مجموعة من الأنماط الحركية المتداخلة وليس نمطا واحدا.
بعض الباحثين حاليا يشطرون مولدات نمط المشي إلى مجموعة كبيرة من مولدات النمط الصغيرة. هم أولا يميزون بين مولدات نمط المشي الخاصة بكل طرف من الأطراف الأربعة. مثلا هم يعتبرون أن هناك مولدات نمط خاصة بالطرف الخلفي الأيمن وأخرى خاصة بالطرف الخلفي الأيسر وهكذا. هذا التمييز يفسر الطابع التناوبي alternating المشاهد في حركة الأطراف أثناء المشي. مثلا لو نظرنا إلى الأطراف الخلفية أثناء المشي فسنجد أن الطرف الأيمن يكون في طور الوقوف أو البسط عندما يكون الطرف الأيسر في طور التأرجح أو الثني، والعكس صحيح. هذا الأمر يمكن تفسيره لو أننا افترضنا أن هناك مولدات نمط منفصلة لكل من الطرفين.
الباحثون وجدوا أن بعض الخلايا الربطية التثبيطية في النصف الأيمن من الحبل الشوكي تصدر أليافا نحو النصف الأيسر من الحبل الشوكي، والعكس صحيح. بناء على ذلك هم يفترضون أن تنشيط مولدات نمط المشي الخاصة بالطرف الأيمن يؤدي إلى تثبيط مولدات نمط المشي الخاصة بالطرف الأيسر، والعكس صحيح. هذه النظرية تفسر الحركة التناوبية للأطراف أثناء المشي.
بعض الباحثين يشطرون مولدات النمط الخاصة بكل طرف من الأطراف إلى مجموعات حسب المفاصل أو الكتل العضلية المختلفة في الطرف. مثلا هم يميزون بين مولد النمط المسؤول عن ثني مفصل الورك، ومولد النمط المسؤول عن بسط مفصل الورك، ومولد النمط المسؤول عن ثني مفصل الركبة، ومولد النمط المسؤول عن بسط مفصل الركبة، إلخ.
وفق هذه الرؤية فإن حركة المشي هي عبارة عن شبكة من المنعكسات الإيقاعية التي تعمل بتناغم فيما بينها، ونفس الأمر ينطبق طبعا على حركة التموج والحركات الإيقاعية الأخرى التي تتولد من الحبل الشوكي والجهاز العصبي المركزي بشكل عام.
الرسم التالي يعبر عن آلية التحكم بالحركة التموجية لدى سمك اللامبري (مصدر الرسم):

الخلايا الموجودة داخل المستطيلين الأسودين تعبر عن مولدات نمط حركة التموج. الخلايا التي تحمل الرمز E هي خلايا ربطية إثارية excitatory interneurons. الخلايا التي تحمل الرمز I هي خلايا ربطية تثبيطية inhibitory interneurons. الخلايا التي تحمل الرمز M هي الخلايا المحركة motor neurons. الخلايا الربطية الإثارية تثير كل أنواع الخلايا الموجودة على نفس جهتها، والخلايا الربطية التثبيطية تثبط كل أنواع الخلايا الموجودة في الجهة المقابلة. الرمز SR-E يعني المستقبلات الحسية الإثارية، والرمز SR-I يعني المستقبلات الحسية التثبيطية. المستقبلات الحسية الإثارية تثير الخلايا الموجودة على نفس جهتها، والمستقبلات الحسية التثبيطية تثبط الخلايا الموجودة على الجهة المقابلة. مولدات النمط تخضع لتأثير ألياف دماغية مفرزة للـ serotonin (اختصاره 5-HT) وألياف دماغية مفرزة للـ GABA وألياف دماغية مفرزة للـ norepinephrine نازلة من الموضع الأزرق locus coeruleus (الألياف الأخيرة غير مبينة على الرسم). الرمز RS يعني خلايا التشكل الشبكي التي تصدر الألياف الشبكية-الشوكية reticulospinal (خلايا إثارية مفرزة للـ glutamate). تنشيط هذه الخلايا يؤدي إلى تنشيط خلايا مولدات النمط في الحبل الشوكي. هذه الخلايا هي واقعة تحت تأثير مركز الحركة الانتقالية في الدماغ الأوسط mesencephalic locomotor region (يحمل الرمز MLR على الرسم) ومركز الحركة الانتقالية في الدماغ البيني diencephalic locomotor region (يحمل الرمز DLR على الرسم). هذان المركزان هما بدورهما خاضعان لتأثير تثبيطي أو كابح من العقد القاعدية basal ganglia.
هذا الرسم التخطيطي يلخص الآلية العصبية المسؤولة عن الحركة الانتقالية لدى سمك اللامبري، الذي هو أحد أكثر الفقاريات بدائية. قشرة المخ المحركة motor cortex لا تظهر في هذا الرسم. السبب هو أن هذه القشرة هي تطور حديث ظهر لأول مرة مع ظهور الثدييات المشيمية placental mammals قبل حوالي 150 مليون عام.
تحكم الدماغ بالحركات العضلية
مما سبق يتبين أمر هام وهو أن الدماغ ليس مسؤولا عن توليد حركة المشي (والركض والقفز والطيران إلخ)، ولكن الدماغ يتحكم بهذه الحركات.
حركة المشي تنتج من تآزر مجموعة من المنعكسات الشوكية. في الحقيقة كل حركات الجسم الفقاري تنتج من تآزر منعكسات شوكية أو بصلية. تنسيق منعكسات المشي وغيرها من المنعكسات يتم أساسا بشكل محلي في القطع العصبية التي تنتج هذه المنعكسات. ما تقوم به المراكز الدماغية العليا هو مؤازرة القطع العصبية الشوكية والبصلية في إنتاج وتنسيق منعكساتها، ولكن المراكز الدماغية العليا لا تتولى بنفسها إنتاج أو تنسيق المنعكسات الشوكية والبصلية.
الآلية المركزية التي تنص على أن تتولى جهة إدارية عليا الإنتاج وتنسيقه هي غير موجودة في عالم الحيوان ولا في الطبيعة بشكل عام. هذه الآلية هي موجودة فقط في بعض المجتمعات البشرية. هذه الآلية لا يمكن أبدا أن تصلح لإدارة أنظمة معقدة كالأنظمة الموجودة في الطبيعة، ولكن كثيرا من البشر لا يفهمون ذلك، لأن القدرات العقلية للبشر محدودة. بعض البشر (الأذكياء) يفهمون حدود وقدرات عقلهم وتفكيرهم، ولكن هناك بشر (أغبياء) لا يفهمون حدود وقدرات عقلهم وتفكيرهم. هؤلاء الأغبياء يميلون دائما إلى إقامة الأنظمة المركزية الصارمة القائمة على منطق تحكم الإدارة العليا بكل صغيرة وكبيرة.
أنا تحدثت عن هذه القضية سابقا وأشرت إلى الحضارة السومرية القديمة التي انهارت بسبب إفراطها في المركزية الإدارية والاقتصادية. الاتحاد السوفييتي هو مثال آخر على فشل الأنظمة المركزية الصارمة. كثير من الناس في زماننا باتوا يفهمون أن الأنظمة الإدارية القائمة على المركزية الصارمة هي أنظمة مناقضة للطبيعة ولا يمكنها أن تنجح، ولكن هناك بعض الناس الذين ما زالوا حتى يومنا هذا يؤمنون بالأنظمة المركزية الصارمة. من هؤلاء مثلا النظام السوري الذي دمر سورية عدة مرات (وليس مرة واحدة) خلال العقود الخمسة الماضية، ولكنه ما زال رغم ذلك يصر على البقاء في السلطة، والمصيبة الأكبر هي أنه يصر على الاستمرار في نفس النهج المركزي التسلطي الفاشل. هذا النظام ما زال يريد أن يتحكم بكل كلمة يتفوه بها كل إنسان في سورية. رئيس هذا النظام (بشار الأسد) ما زال يريد أن يفكر بنفسه عوضا عن الـ 23 مليون إنسان في سورية.
أعتذر عن خلط السياسة بموضوع بعيد عن السياسة.
العلاقة بين الدماغ والحركة الانتقالية هي موضوع إشكالي.
الحركة الانتقالية بواسطة الأطراف الممفصلة تتم عبر تقليص عدد هائل من الألياف العضلية بواسطة عدد هائل من الألياف العصبية المحركة. مثلا لو نظرنا إلى الرِجلين فسنجد أنهما تحويان عددا هائلا من الألياف العضلية، وهناك أيضا عظام متعددة ومفاصل متعددة في كل من الرجلين. التحكم بالأطراف يبدو أمرا معقدا للغاية. وهناك مشكلة إضافية أشار إليها في النصف الأول من القرن العشرين باحث روسي اسمه Nikolai Bernstein. هذه المشكلة هي مشكلة درجات الحرية degrees of freedom problem في التحكم العضلي.
المقصود بمشكلة درجات الحرية هو ما يلي باختصار: الأطراف الأربعة تحوي عددا هائلا من الألياف العضلية والأوتار والعظام والمفاصل، وهذا العدد الهائل هو زائد عن الحاجة وليس ضروريا لتحريك الأطراف.
لنفرض أنني أريد أن أرسل شخصا إلى المتجر لكي يشتري لي علبة من العصير، ولنفرض أنني لا أستطيع أن أرسل شخصا واحدا فقط ولكنني مضطر لإرسال ألف شخص في آن واحد لكي ينفذوا هذه المهمة. عندما أريد أن أرسل هؤلاء الأشخاص الألف فإنني مضطر لإعطائهم تعليمات. المشكلة هي أن المهمة المطلوبة منهم هي مهمة تافهة ولا تستحق إعطاء تعليمات لألف شخص. لو أنني أعطيت التعليمات لشخص واحد فقط (أحضر لي العصير من المتجر) فإن هذا سيفي بالغرض، ولكنني مضطر لإرسالهم جميعا، وبالتالي لا بد أن يحصلوا جميعا على تعليمات.
التعليمات التي سأعطيها لهم هي مجرد مضيعة للوقت والجهد وليس لها فائدة عملية. يكفي أن أقول لشخص واحد منهم “أحضر لي العصير من المتجر” حتى أضمن تنفيذ المهمة، وأما الكلام الذي سأقوله للأشخاص الـ 999 الآخرين فهو مجرد كلام زائد.
هذا المثال يشبه مشكلة “درجات الحرية” التي طرحها Nikolai Bernstein. من الناحية النظرية الإنسان يمكنه أن يحرك أطرافه بواسطة عدد قليل من الإشارات العصبية، ولكننا نعلم أن هناك في الأطراف عددا هائلا من الألياف العضلية التي تستقبل عددا هائلا من الألياف العصبية. كيف يمكن للدماغ البشري أن يتعامل مع هذا العدد الزائد من الألياف العضلية والألياف العصبية؟
هناك نظريات عديدة طرحت للإجابة على مشكلة “درجات الحرية”. Bernstein نفسه طرح نظرية “تآزر العمل العضلي” muscle synergies. حسب هذه النظرية فإن كل إشارة عصبية تخرج من الدماغ تذهب إلى عدد كبير من الألياف العضلية وليس إلى ليف واحد، وهذا يوفر الجهد على الدماغ ويقلل من “درجات الحرية” الفائضة.
نظرية “تآزر العمل العضلي” لم تتوافق مع بعض التجارب العملية. في الآونة الأخيرة طرحت نظرية جديدة تبدو أفضل. النظرية الجديدة تنص على أن الدماغ لا يرسل إشارات عصبية نحو كل الألياف العضلية ولكنه يرسل إشارات عصبية نحو بعض الألياف العضلية. مثلا لنفرض أنني أقف أمام باب وأنني أريد أن أمد يدي نحو مقبض الباب. عندما أبدأ في مد يدي نحو مقبض الباب فإن دماغي يرسل إشارات عصبية نحو بعض الألياف العضلية في ذراعي وليس كل الألياف. الألياف التي يرسل دماغي الإشارات إليها هي الألياف المهمة التي من شأن التأثير فيها أن يحسن من كفاءة الحركة الإجمالية.
لنعد مجددا إلى مثال العصير. Bernstein افترض أنني يجب أن أعطي التعليمات لكل شخص من الأشخاص الألف الذين أرسلتهم لشراء العصير، ولكن في الحقيقة أنا لست مضطرا لإعطائهم التعليمات جميعا. لو أنني كنت أحكم دولة مركزية صارمة (على غرار الاتحاد السوفييتي الذي عاش فيه Bernstein) فإنني كنت سأضطر لإعطاء التعليمات لكل شخص من الأشخاص الألف، ولكنني لا أحكم دولة مركزية صارمة، وبالتالي أنا لست مضطرا لأن أعطي التعليمات للأشخاص الألف. ما سأفعله هو ما يلي: أنا سأستدعي شخصا واحدا أعلم (من تجاربي السابقة) أنه الأقدر على إحضار العصير، وسأطلب من هذا الشخص أن يحضر لي العصير، وبالنسبة للأشخاص الـ 999 الآخرين فأنا لن أقول لهم شيئا وسأتركهم يتصرفون على سجيتهم، لأن تصرفاتهم لن تؤثر على نجاح المهمة. هذا المثال يعبر ربما عن الطريقة التي يتعامل بها الدماغ مع الحركات العضلية.
تنسيق حركة الأطراف لا يحتاج لأوامر من الدماغ، لأن الحبل الشوكي يحوي منعكسات قادرة على تنسيق حركة الأطراف محليا ودون تدخل من المراكز العليا. ما يقوم به الدماغ هو أنه يتحكم ببعض عضلات الأطراف بهدف زيادة تنسيق الحركة أو تحسينها. الدماغ لا يولد الحركة ولا ينسقها ولكنه يحسنها، وهو لا يتدخل في عمل جميع الألياف العضلية ولكنه يتدخل في عمل جزء منها.
كل ليف عضلي هو مهم بالنسبة لحركة معينة. الدماغ يحفظ الألياف العضلية المهمة بالنسبة لكل حركة من الحركات، وعندما يقرر الدماغ تنفيذ حركة معينة فإنه يرسل إشارات عصبية نحو الألياف المهمة بالنسبة لتلك الحركة.
لماذا يثق بعض الزعماء بالنساء أكثر من الرجال؟

أنا لا أعرف شيئا عن شخصية الرئيس السوري بشار الأسد، لأنني لم أقابله في حياتي ولم أعاشره حتى أقيم شخصيته، ولكنني قرأت الكثير من المقالات والكتابات التي تتحدث عن شخصيته.
الدماغ البشري (11)
كثير من الكائنات الحية تملك القدرة على الحركة. بعض الكائنات الحية تستطيع أن تحرك أجزاء من جسمها، وبعضها تستطيع أن تنقل جسمها بكامله من مكان إلى آخر. الحركة التي تؤدي لنقل الجسم بكامله من مكان إلى آخر تسمى الحركة الانتقالية locomotion.الحركة الانتقالية هي خاصية بيولوجية بدائية جدا، بدليل انتشارها الواسع لدى الكائنات الحية أحادية الخلية حقيقية النواة eukaryotes.الباحث السويدي Carolus Linnaeus (في القرن 18) قسم الكائنات الحية إلى مملكتين:
- مملكة الحيوان Regnum Animale
- مملكة النبات Regnum Vegetabile
هذا التقسيم لم يفرد مملكة خاصة للكائنات الحية المجهرية أحادية الخلية التي اكتشفها الهولندي Antonie van Leeuwenhoek في نهاية القرن 17. باحثو البيولوجيا ظلوا حتى القرن 19 يصنفون الكائنات الحية أحادية الخلية ضمن المملكتين الحيوانية والنباتية.
في القرن 19 ظهر المصطلحان التاليان:
- الحيوانات البدائية protozoans (مشتق من كلمة (zō̃ion) ζῷον التي تعني “حيوان”)
- النباتات البدائية protophytes (مشتق من كلمة (phytόn) φυτόν التي تعني “نبات”)
هذان المصطلحان يغطيان الكائنات الحية أحادية الخلية. “الحيوانات البدائية” protozoans هي تسمية أطلقت على الكائنات أحادية الخلية التي “تشبه الحيوانات” (من حيث امتلاكها القدرة على الحركةmotility ، ومن حيث اعتمادها على الكائنات الحية الأخرى في الغذاء heterotrophy). “النباتات البدائية” protophytes هي تسمية أطلقت على الكائنات أحادية الخلية التي “تشبه النباتات” (من حيث قدرتها على صنع غذائها بنفسها autotrophy عبر التركيب الضوئي photosynthesis).
في أواسط القرن 19 كثير من الباحثين صاروا يشعرون أن تقسيم الكائنات أحادية الخلية إلى حيوانات ونباتات هو تقسيم اعتباطي وغير علمي: هناك كائنات أحادية الخلية لا يمكن تصنيفها على أنها حيوان أو نبات. لهذا السبب اقترح الباحث الألمانيErnst Haeckel في عام 1866 إنشاء مملكة جديدة هي “مملكة الطلائعيات” Regnum Protista (المعنى الحرفي لكلمة protista هو “الأكثر بدائية”، ولكنني ترجمتها إلى “طلائعيات” لأن هذه هي الترجمة الشائعة). مملكة الطلائعيات كانت تضم “الكائنات المحايدة”، أي الكائنات التي ليست حيوانات ولا نباتات. لاحقا Haeckel عدّل تعريف هذه المملكة بحيث صارت تضم كل الكائنات وحيدة الخلية.
خلاصة تصنيفHaeckel هي ما يلي:
- مملكة الطلائعيات Regnum Protista (الكائنات وحيدة الخلية unicellular organisms)
- مملكة النباتات Regnum Plantae
- مملكة الحيوانات Regnum Animalia
في القرن العشرين ظهر اختراع مهم هو الميكروسكوب الإلكتروني electron microscope. هذا الميكروسكوب كشف أن الكائنات أحادية الخلية هي في الحقيقة صنفان مختلفان:
- كائنات بدائية النواة prokaryotes
- كائنات حقيقية النواة eukaryotes

الكائنات “حقيقية النواة” لها تركيب مشابه لتركيب الخلايا الحيوانية والنباتية، وأما الكائنات “بدائية النواة” فهي تتميز بتركيب أكثر بدائية. لهذا السبب الباحثون في أواسط القرن العشرين صاروا يرون أن الكائنات بدائية النواة هي الأقدم تطوريا. حسب النظرة الجديدة فإن الكائنات حقيقية النواة تطورت من الكائنات بدائية النواة، والكائنات متعددة الخلايا multicellular organisms (الحيوانات والنباتات) تطورت من الكائنات حقيقية النواة.
بناء ذلك ظهر تصنيف جديد يقسم الكائنات الحية إلى مجموعتين كبيرتين:
- إمبراطورية بدائيات النواة Imperium Prokaryota
- إمبراطورية حقيقيات النواة Imperium Eukaryota
الإمبراطورية الثانية تنقسم إلى ثلاث ممالك هي الطلائعيات والنباتات والحيوانات، وأما الإمبراطورية الأولى فهي أنتجت مملكة واحدة هي “مملكة الأحاديات” Regnum Monera.
مملكة النباتات حسب التصنيف التقليدي تضم الفطريات Fungi، ولكن هذا التصنيف هو تصنيف اعتباطي وليس تصنيفا علميا. Ernst Haeckel أدرك ذلك، ولهذا السبب هو أخرج الفطريات من مملكة النباتات وألحقها بمملكة الطلائعيات، ولكن معاصريه تجاهلوا ذلك وظلوا يصنفون الفطريات مع النباتات. في عام 1969 اقترح الباحث الأميركي Robert Whittaker إنشاء مملكة جديدة ضمن إمبراطورية حقيقيات النواة هي “مملكة الفطريات” Regnum Fungi. هذا الاقتراح لقي قبولا واسعا (على الأقل في أميركا) وبناء عليه صار تصنيف الكائنات الحية هو كما يلي:
- إمبراطورية بدائيات النواة Imperium Prokaryota
- مملكة الأحاديات Regnum Monera
- إمبراطورية حقيقيات النواة Imperium Eukaryota
- مملكة الطلائعيات Regnum Protista
- مملكة الفطريات Regnum Fungi
- مملكة النباتات Regnum Plantae
- مملكة الحيوانات Regnum Animalia
خلال العقود الأخيرة من القرن العشرين برز دور الدراسات الجزيئية والجينية. هذه الدراسات دمرت تماما التصنيفات التقليدية التي سردناها في الأعلى.
أول نتيجة للدراسات الجزيئية هي تقسيم الكائنات بدائية النواة prokaryotes إلى مجموعتين مستقلتين:
- البكتيريا Bacteria
- العتيقات Archaea

من غير المعروف أي المجموعتين هي الأقدم، ومن غير المعروف ما هي العلاقة بين هاتين المجموعتين وبين مجموعة حقيقيات النواة eukaryotes. الشيء الوحيد الذي يتفق عليه كثير من الباحثين حاليا هو أن الكائنات الحية تتوزع ضمن المجموعات الثلاث الظاهرة في الرسم التالي:

كل مجموعة من هذه المجموعات تسمى domain. المعنى الحرفي لهذه الكلمة هو “أرض سيّد” (land) belonging to a lord.
هناك نظرية ترى أن الأراضي الثلاث (bacteria و archaea و eukaryota) هي متحدرة من أصل مشترك كان موجودا في العصر السابق لظهور الخلايا.
الخلايا cells هي بنى معقدة، والباحثون لا يعتقدون أن الخلايا هي أقدم أشكال الحياة. هم يرون أن هناك أشكالا حية أكثر بدائية من الخلايا كانت موجودة قبل ظهور الخلايا، ولكن معلوماتهم عن هذه الأشكال الحية البدائية هي محدودة للغاية.
بالنسبة لمجموعة حقيقيات النواة eukaryotes فهناك حاليا جدل كبير حول تصنيف الكائنات المنضوية تحتها.
بعض الدراسات الحديثة تقسم حقيقيات النواة إلى مجموعتين كبيرتين:
- ثنائيات المجداف bikonts
- أحاديات المجداف unikonts
كلمة (kontós) κοντός تعني بالإنكليزية punting-pole (انظر القاموس). معنى punting-pole هو المجداف الذي يستخدم لتحريك قارب شبيه بالطوف يسمى punt (هذه صورته). إذن كلمتا bikonts و unikonts تعنيان حرفيا “ثنائيات المجداف” و”أحاديات المجداف”. المقصود بثنائيات المجداف هو الخلايا التي تملك سوطين two flagella، والمقصود بأحاديات المجداف هو الخلايا التي تملك سوطا واحدا one flagellum.
تصنيف ثنائيات المجداف يضم النباتات، وتصنيف أحاديات المجداف يضم الفطريات والحيوانات. بالنسبة للكائنات وحيدة الخلية فهي تتوزع بين التصنيفين.
بعض الدراسات ترى أن الكائنات ثنائية المجداف هي الأقدم تطوريا، وأن الكائنات أحادية المجداف هي مشتقة من كائن ثنائي المجداف.
كثير من الدراسات ترى أن الكائنات الأكثر بدائية بين ثنائيات المجداف (وبالتالي بين حقيقيات النواة) هي الكائنات المسماة excavates. هذا التصنيف يضم الكائن المسمى Euglena بالإضافة إلى عدد من الطفيليات المشهورة طبيا من قبيل Trypanosoma و Naegleria و Giardia و Trichomonas.

حسب بعض الدراسات فإن أقرب الكائنات إلى الـ excavates هي النباتات. هذا يوحي ربما بأن النباتات هي أكثر بدائية من الحيوانات والفطريات.
تصنيف النباتات حاليا هو محل جدل. بعض الدراسات تشطر المملكة التي كانت معروفة سابقا باسم مملكة النباتات إلى عدد كبير من المجموعات المستقلة، وبعض الدراسات تشطر مملكة النباتات إلى مجموعتين فقط:
- chromalveolates (“المجوّفات الصبغية”)
- archaeplastidans (“البلاستيديات العتيقة”)
المجموعة الثانية (archaeplastidans) تضم النباتات الحقيقية، وأما المجموعة الأولى فهي تضم أجناسا من الطحالب.
مجموعة الكائنات أحادية المجداف unikonts تقسم إلى مجموعتين:
- الحيوانات الأميبية amoebozoans
- خلفيات المجداف opisthokonts
مجموعة “الحيوانات الأميبية” amoebozoans هي جزء من المجموعة التي كانت تسمى سابقا “الأميبا” amoeba أو “شبيهات الأميبا” amoeboids. الجزء الآخر من مجموعة الأميبا يسمى rhizarians وهو ينتمي لمجموعة chromalveolates التي ذكرناها في الأعلى.

خلفيات المجداف opisthokonts هي المجموعة التي أنتجت الفطريات والحيوانات. بعض الدراسات تشطر هذه المجموعة إلى شطرين: شطر أنتج الفطريات وشطر آخر أنتج الحيوانات. الشطر الذي أنتج الحيوانات يسمى أحيانا holozoans (“الحيوانات الكاملة”). هذا الشطر أنتج مجموعة من الكائنات تسمى filozoans (“الحيوانات الخيطية”)، وهذه بدورها أنتجت مملكة الحيوانات التي تسمى أيضا metazoans (“الحيوانات التالية”).
أكثر الكائنات بدائية ضمن مملكة الحيوانات هي الإسفنجيات poriferans و”الحيوانات الصفيحية” placozoans. هذه الكائنات تصنف تحت عنوان “جانب الحيوانات” parazoans. بقية أعضاء مملكة الحيوانات تصنف تحت عنوان “الحيوانات التالية الحقيقية” eumetazoans.


الحيوانات التالية الحقيقية eumetazoans تنقسم إلى “الشعاعيات” radiates وثنائيات الجانب bilaterians.
الشعاعيات تضم المِشْطِيات ctenophores واللاسعات cnidarians. أجسام هذه الكائنات تتميز بالتناظر الشعاعي radial symmetry، بخلاف أجسام ثنائيات الجانب التي تتميز بالتناظر ثنائي الجانب bilateral symmetry. الفرق بين هذين النوعين من التناظر هو مذكور في هذا المقال.



مجموعة الحيوانات ثنائية الجانب bilaterians تضم عددا كبيرا جدا من الفروع. معظم هذه الفروع تدخل تحت مسمى الديدان worms (هذا المسمى هو مجرد تصنيف شعبي وليس تصنيفا علميا). إلى جانب الديدان مجموعة ثنائيات الجانب تضم أيضا مفصليات الأرجل arthropods وشوكيات الجلد echinoderms والحبليات chordates.

أبسط أنواع الحبليات chordates هي ربما “الحبليات الرأسية” cephalochordates المتمثلة بحيوان lancelet أو amphioxus (تحدثنا عنه في مقالات سابقة). الفقاريات vertebrates هي فرع من الحبليات. أنا تحدثت عن تطور الفقاريات في هذا المقال.

الحركة الانتقالية لدى الكائنات وحيدة الخلية
ما سبق هو عرض موجز لتطور الحيوانات. من هذا العرض يتبين أن الحيوانات هي فرع من حقيقيات النواة eukaryotes. اللافت هو أن خاصية الحركة motility هي على ما يبدو خاصية عتيقة جدا في حقيقيات النواة، بدليل أن الباحثين حاليا يصنفون حقيقيات النواة حسب عدد أسواطها إلى ثنائيات مجداف bikonts وأحاديات مجداف unikonts. حسب علمي فإن الوظيفة الأساسية للأسواط flagella هي الحركة الانتقالية locomotion. هذه الوظيفة تظهر واضحة لدى حقيقيات النواة البدائية المسماة excavates. هذه الكائنات تستخدم الأسواط للحركة الانتقالية.
بعض حقيقيات النواة تستخدم وسيلة أخرى للحركة الانتقالية هي الأهداب المتحركة motile cilia. الفرق بين عمل الأهداب المتحركة وبين عمل الأسواط هو طفيف.

هناك طريقة ثالثة للحركة الانتقالية لدى حقيقيات النواة هي “الأقدام الزائفة” pseudopodia التي تستخدمها الأميبا amoeba.

من الناحية التطورية الأسواطflagella هي ربما الأقدم. الأسواط هي موجودة لدى بدائيات النواة prokaryotes، ولكن الأهداب cilia والأقدام الزائفة pseudopodia هي غير موجودة لدى بدائيات النواة. أيضا لو نظرنا في مجموعة حقيقيات النواة فسنرى أن الـ excavates (التي هي الأكثر بدائية بين حقيقيات النواة) تعتمد على الأسواط وليس الأهداب أو الأقدام الزائفة.
بقايا الحركة السوطية والهدبية والقدمية الزائفة لدى البشر
أنماط الحركة التي سردناها في الأعلى ما زالت موجودة في أجسام البشر. بالنسبة للحركة عبر الأسواط فهي مثلا الطريقة التي يتحرك بواسطتها الحيوان المنوي sperm:

الحركة الهدبية هي موجودة لدى البشر في الخلايا ذات الأهداب المبطنة للقنوات التنفسية والتناسلية. حركة أهداب هذه الخلايا لا تؤدي لتحريك الخلايا ولكنها تؤدي لتحريك المفرزات الموجودة داخل القنوات التنفسية والتناسلية.

الحركة عبر الأقدام الزائفة هي الطريقة التي تتحرك بها خلايا الدم البيضاء:

الحركة الانتقالية لدى ثنائيات الجانب
بعض الحيوانات ثنائية الجانب ما زالت تعتمد على الأهداب المتحركة في حركتها الانتقالية. مثلا الديدان المسطحة platyhelminthes تعتمد على الأهداب المتحركة في حركتها الانتقالية، ولهذا السبب هي تتحرك ببطء شديد.

كثير من الديدان الأخرى تعتمد على نمط من الحركة يسمى التموّج undulation. هذا النمط من الحركة هو المشاهد لدى الديدان السهمية arrow worms والديدان المدورة roundworms وغيرها. هذا النمط من الحركة هو المشاهد أيضا لدى الحبليات والفقاريات البدائية.

تطور الحركة لدى ثنائيات الجانب هو مرتبط بظهور خاصية “التقطع المتتالي” metamerism في أجسامها.
بعض النظريات ترى أن التقطع المتتالي إنما ظهر بهدف تحسين الحركة. الجسم غير المقطع تقطيعا متتاليا هو عمليا أشبه بكتلة صخرية، لأن جوف الجسم coelom هو مملوء بالماء، والماء ينقل الضغط ويوزعه بشكل متساوي في جميع أنحاء الجسم. لو تقلصت إحدى عضلات جدار الجسم فستنتقل القوة الناتجة عن هذا التقلص عبر ماء الجوف نحو جميع مناطق الجسم. هذا الوضع يجعل من الصعب توجيه قوة التقلص العضلي نحو جزء محدد من الجسم، بالتالي الحيوان لا يمكنه أن يحرك جزءا محددا من جسمه ولكنه مضطر دائما للتحرك ككتلة واحدة.
الجسم المقطع إلى قطع متتالية metameres لا يعاني من هذه الصعوبة، لأن الحواجز التي تفصل بين القطع المتتالية تمنع انتقال الضغط عبر ماء الجوف من قطعة متتالية إلى أخرى. بالتالي الحيوان يمكنه أن يحرك كل قطعة متتالية على حدة. هذا يجعل جسم الحيوان مفصّلا hinged (بمعنى أنه يحوي مفاصل)، وبالتالي يحسن قدرة الحيوان على تحريك جسمه.
إذن التقطع المتتالي يعطي الحيوان القدرة على تنفيذ الحركات بدقة أكبر، ولكنه في نفس الوقت يخلق مشكلة جديدة هي ضرورة التنسيق الحركي motor coordination بين القطع المتتالية. تحريك قطعة واحدة لا يحتاج لتنسيق، ولكن تحريك عدد كبير من القطع هو بحاجة لتنسيق حتى لا تتعارض حركات القطع مع بعضها. تنسيق حركات القطع المتتالية يتم عبر الجهاز العصبي المركزي. هذا الجهاز يتكون لدى الديدان الحلقية annelids من حبلين بطنيين ventral cords وعقدتين دماغيتين cerebral ganglia. تكوين الجهاز العصبي المركزي لدى مفصليات الأرجل arthropods والحبليات chordates هو شبيه بتكوين الجهاز العصبي المركزي لدى الديدان الحلقية (كما بينا في مقالات سابقة).
الحركة التمعجية
معظم الديدان تعتمد على التموج undulation في حركتها الانتقالية، ولكن دودة الأرض earthworm (من الديدان الحلقية) طورت نوعا مميزا من الحركة الانتقالية يسمى peristalsis (“التَّمَعُّج”). المقطع التالي يبين هذه الحركة:
حركة “التمعج” هي موجودة لدى البشر ضمن القناة الهضمية، خاصة في المريء والأمعاء. حركة المريء والأمعاء لدى البشر تشبه حركة دودة الأرض الظاهرة في المقطع أعلاه، ولكنها ليست حركة انتقالية وإنما حركة تهدف لنقل الغذاء عبر القناة الهضمية.
الحركة التموجية لدى الفقاريات
الحركة التموجية undulation هي شائعة لدى ثنائيات الجانب البدائية. كثير من أنواع الديدان تتحرك بهذه الطريقة، وأيضا الحبليات والفقاريات البدائية تتحرك بهذه الطريقة.
المقصود بـ”الفقاريات البدائية” هو الأسماك، وخاصة الأسماك بلا فكين jawless fish كسمكة اللامبري lamprey. ما يلي مقطع يظهر حركة التموج لدى سمكة اللامبري:
حركة التموج الانتقالية هي موجودة أيضا لدى الأفاعي:
الأفاعي هي من الزواحف، ولكنها تعرضت لطفرة جينية جعلتها تفقد أطرافها limbs. لهذا السبب هي ارتدت نحو الحركة التموجية المميزة للفقاريات البدائية.
بقايا الحركة التموجية لدى الإنسان
في مقال سابق تحدثت عن بقايا الحركة التموجية لدى الإنسان. عضلات الجسم المسؤولة عن تنفيذ الحركة التموجية هي العضلات الباسطة المحورية axial extensor muscles. هذه العضلات هي مسؤولة عن بسط أو ثني العمود الفقري، وبالتالي هي مسؤولة عن الحركة التموجية. في الدماغ البشري هناك أنوية مسؤولة عن التحكم بهذه العضلات تقع في الدماغ الخلفي (hindbrain) rhombencephalon، وتحديدا في التشكل الشبكي reticular formation. هذه الأنوية هي مسؤولة لدى البشر عن عدد من المنعكسات البدائية التي تظهر خلال مرحلة الطفولة الباكرة. هذه المنعكسات ليس لها فائدة بالنسبة للبشر، ولهذا السبب يتم تثبيطها بشكل نهائي خلال مرحلة الطفولة، ولكن من الممكن أن هذه المنعكسات هي بقايا تطورية من زمن الفقاريات الباكرة، أي أن لها علاقة بالحركة الانتقالية التموجية.
الأنوية المسؤولة عن تقليص وإرخاء العضلات الباسطة المحورية (والتي كانت في السابق مسؤولة عن الحركة التموجية) اكتسبت لدى البشر وظيفة جديدة هي التحكم بالتوتر العضلي muscle tone للعضلات الباسطة المحورية. التوتر العضلي للعضلات الباسطة المحورية هو ضروري للمحافظة على استقرار الهيكل العظمي ومنع السقوط؛ لهذا السبب العضلات الباسطة المحورية تسمى لدى البشر باسم “العضلات المضادة للجاذبية” anti-gravity muscles.
الحركة بواسطة الأطراف الممفصلة
الحركة الانتقالية بواسطة الأطراف الممفصلة jointed limbs (الأطراف ذات المفاصل) هي شكل متطور من الحركة الانتقالية يميز نوعين من الحيوانات التي تعيش خارج الماء هما مفصليات الأرجل arthropods ورباعيات الأرجل tetrapods (رباعيات الأرجل هي فرع من الفقاريات يشمل الفقاريات التي تملك أربعة أطراف).
الحركة الانتقالية بواسطة الأطراف الممفصلة تشمل حركات المشي والركض والقفز والطيران بالأجنحة.
هذا الشكل من الحركة الانتقالية يتطلب وجود الأطراف الممفصلة ويتطلب القدرة على التحكم بها بشكل إيقاعي.
النواة الحمراء
أشرت في مقال سابق إلى النواة الحمراء red nucleus (باللاتينية: nucleus ruber). هذه النواة تظهر في مقطع الدماغ الأوسط المنقاري rostral midbrain عند مستوى التليلتين العلويتين superior colliculi:

وظيفة النواة الحمراء مجهولة، ولكن هناك بعض النظريات حولها.
النواة الحمراء هي في الحقيقة نواتان اثنتان:
- النواة الحمراء كبيرة الخلايا magnocellular red nucleus (اختصارا RNm)
- النواة الحمراء صغيرة الخلايا parvocellular red nucleus (اختصارا RNp)
النواة الحمراء كبيرة الخلايا RNm تمثل القسم الذيلي من النواة الحمراء، والنواة الحمراء صغيرة الخلايا RNp تمثل القسم المنقاري من النواة الحمراء.
بعض الطروحات الحديثة ترى أن الوظيفة الأصلية للنواة الحمراء كبيرة الخلايا RNm هي تحريك عضلات الأطراف.
Although the RN can be differentiated in some fishes and limbless reptiles, a rubrospinal tract has so far been demonstrated only in rays which use their pectoral fins for locomotion [1]. The fins of the ray are likely precursors to the limbs of terrestrial vertebrates [7]. Rubrospinal tracts are also absent in boid snakes and sharks who use axial movements for locomotion rather than limbs [9]. Such data further implicates RNm and the corresponding rubrospinal tract in limb movement.
النواة الحمراء كبيرة الخلايا RNm هي مصدر لطريق عصبي نازل نحو الحبل الشوكي يسمى “الطريق الأحمر-الشوكي” rubrospinal tract. هذا الطريق هو غير موجود لدى “الفقاريات السفلى” (أي الأسماك)، باستثناء نوع من الأسماك الغضروفية يسمى rays (الاسم العلمي batoids). هذا النوع من الأسماك يتميز عن غيره بأنه يستخدم زعانفه للحركة الانتقالية locomotion.
الصور التالية تبين سمك الـ ray:


هناك اعتقاد بأن زعانف سمك الـ ray لها نفس أصل أطراف الفقاريات رباعية الأرجل.
الطريق الأحمر-الشوكي هو موجود لدى الزواحف باستثناء الأفاعي (التي تفتقد للأطراف).
الملاحظات السابقة تدعم الرأي القائل بأن وظيفة الطريق الأحمر-الشوكي هي تحريك الأطراف. هذا الطريق هو موجود حصرا لدى الفقاريات التي تتحرك بواسطة الأطراف.
النواة الحمراء صغيرة الخلايا RNp هي على ما يبدو غائبة لدى الأسماك والبرمائيات، وهي هامشية لدى بقية الفقاريات باستثناء الرئيسيات primates. اللافت هو أن نمو الـ RNp لدى الرئيسيات ترافق مع اضمحلال وضمور الـ RNm والطريق الأحمر-الشوكي النازل منها.
Studies of the mammalian red nucleus demonstrate that larger cells are found in RNm, whereas medium and small cells are typically found in RNp [3]. The volume of the nucleus taken up by these parts becomes reversed over the course of time and across species. In reptiles and birds the nucleus is composed almost entirely of large cells [4]. This large-celled RNm is the most likely source of the contralateral projections of the rubrospinal tract prominent in early vertebrates [1]. Yet as the rubrospinal tract has regressed in primates so has the histological presence of RNm. The number of large cells in the RN seems to diminish progressively as one passes from carnivores to baboons to anthropoids to man [2]. Morphological studies in humans show an increased development of the RNp – enough that RNm and RNp are completely independent of each other in humans [5].
[…]
The rubrospinal pathway and its cell bodies in RNm have progressively declined from lower mammals to primates [4]. Some researchers note that the RNm, initially involved in the automatic execution of learned motor skills, has been progressively replaced by the pyramidal system [1].
النواة الحمراء كبيرة الخلايا RNm اضمحلت بشكل تدريجي لدى الرئيسيات حتى وصلت إلى أقصى درجات اضمحلالها لدى البشر. الطريق الأحمر-الشوكي لدى البشر يصل فقط نحو القسم الرقبي cervical من الحبل الشوكي الذي يتحكم بالطرفين العلويين (الذراعان)، ولكنه لا يصل نحو القسم القَطَني lumbar من الحبل الشوكي الذي يتحكم بالطرفين السفليين (الرجلان). لدى بقية الثدييات (مثلا القطط) الطريق الأحمر-الشوكي يصل نحو القسم القطني من الحبل الشوكي.
في المقابل النواة الحمراء صغيرة الخلايا RNp نمت بشكل تدريجي لدى الرئيسيات حتى وصلت إلى أقصى درجات نموها لدى البشر. هذه النواة هي مصدر ألياف الطريق الأحمر-الزيتوني rubro-olivary tract الذي يصل نحو الزيتونة السفلية inferior olive.
الـ RNp تتلقى أليافا نازلة من قشرة المخ الحسية والمحركة والقشرة أمام الجبهية prefrontal (المسؤولة عن المعالجة المنطقية أو ما يسمى “الوظائف التنفيذية” executive functions). الـ RNp تتلقى أيضا أليافا من النواة المسننة في المخيخ dentate nucleus. اتصالات الـ RNp تشبه اتصالات قشرة المخ المحركة، ولهذا السبب بعض الباحثين يرون أن الـ RNp هي جزء من منظومة “الجهاز الهرمي” pyramidal system. هم يرون أن نمو الـ RNp ترافق مع نمو الجهاز الهرمي لدى الرئيسيات وهيمنة هذا الجهاز على “الجهاز خارج الهرمي” extrapyramidal system.
المقصود بالجهاز الهرمي هو المنظومة التحريكية التي تديرها قشرة المخ (المنظومة التحريكية الواعية)، والمقصود بالجهاز خارج الهرمي هو المنظومة التحريكية التي يديرها جذع الدماغ (المنظومة التحريكية غير الواعية). نحن عددنا طرق الجهاز الهرمي في مقالات سابقة (منها مثلا الطريق القشري-الشوكي corticospinal tract والطريق القشري-البصلي corticobulbar tract). أيضا نحن تحدثنا عن بعض طرق الجهاز خارج الهرمي في مقالات سابقة (منها الطريق الشبكي-الشوكي reticulospinal tract والطريق الدهليزي-الشوكي vestibulospinal tract).
النواة الحمراء كبيرة الخلايا RNm تستقبل أليافا نازلة من قشرة المخ المحركة، وتستقبل أليافا صاعدة من الحبل الشوكي تسمى الطريق الشوكي-الأحمر spinorubral tract. هذا الطريق يشبه الطريق الشوكي-الشبكي spinoreticular tract والطريق الشوكي-الدهليزي spinovestibular tract. اتصالات الـ RNm تتناسب مع كونها جزءا من الجهاز خارج الهرمي extrapyramidal system.
الـ RNm تتلقى أليافا من نواة مخيخية تسمى “النواة المتوسطة” interposed nucleus. على الأغلب أن وظيفة هذه النواة لها علاقة بتحريك الأطراف. النواة المسننة dentate nucleus (التي ترسل أليافا نحو الـ RNp ونحو قشرة المخ المحركة) هي مرتبطة بالجهاز الهرمي. هناك نواة مخيخية ثالثة هي “النواة الأَوْجِية” fastigial nucleus. هذه النواة هي أقدم الأنوية المخيخية وهي ترسل أليافا نحو الأنوية الدهليزية. على الأغلب أن وظيفة النواة الأوجية لها علاقة بتحريك العضلات الباسطة المحورية للجسم (تشرف على تنظيم وضعية الجسم posture عبر التحكم بالتوتر العضلي muscle tone).
الوضعية مزالة القشرة
هناك حالة طبية تسمى “الوضعية مزالة الدماغ” decerebrate posture. هذه الوضعية تظهر لدى الأشخاص المصابين بأذية كبيرة في جذع الدماغ تحت مستوى النواة الحمراء.

في هذه الوضعية هناك تقلص عام للعضلات الباسطة المحورية (وهو ما يؤدي إلى انبساط أو انتصاب الجسم).
ما هو سبب هذه الوضعية؟
السبب هو انقطاع الألياف النازلة من قشرة المخ نحو التشكل الشبكي والأنوية الدهليزية. هذه الألياف تمارس تأثيرا مثبطا أو منظما للأنوية الشبكية والدهليزية المسؤولة عن تقليص العضلات الباسطة المحورية (عبر الطريق الشبكي-الشوكي reticulospinal tract والطريق الدهليزي-الشوكي vestibulospinal tract).
هناك وضعية أخرى ملفتة أكثر من الوضعية سالفة الذكر تسمى “الوضعية مزالة القشرة” decorticate posture. هذه الوضعية تظهر لدى الأشخاص المصابين بأذية فوق مستوى النواة الحمراء.

هذه الوضعية تشبه وضعية المومياء المصرية. في هذه الوضعية هناك انبساط في عضلات الجسم ما عدا عضلات الطرفيين العلويين التي هي في حالة انقباض أو ثني.
تفسير هذه الوضعية هو أن الأذية فوق مستوى النواة الحمراء تزيل التثبيط القشري عن النواة الحمراء كبيرة الخلايا RNm، وهذا يؤدي إلى تنشيط الطريق الأحمر-الشوكي rubrospinal tract الذي يؤثر في الطرفين العلويين دون الطرفين السفليين (لأن أليافه لدى البشر لا تنزل نحو القسم القطني lumbar من الحبل الشوكي).
الوضعية مزالة القشرةdecorticate توحي بأن الطريق الأحمر-الشوكي يمارس تأثيرا مقبضا لعضلات الأطراف. هذا التأثير يهدف في الأصل لتحريك الأطراف.
وظيفة النواة الحمراء كبيرة الخلايا
في الأعلى قلنا أن وظيفة النواة الحمراء كبيرة الخلايا RNm هي على الأغلب تحريك الأطراف بهدف الحركة الانتقالية locomotion. نشوء هذه النواة هو مرتبط بظهور الحركة الانتقالية عبر تحريك الأطراف.
السؤال هو لماذا ضمرت هذه النواة لدى الرئيسيات عموما والبشر خصوصا؟
الميزة الأساسية التي تميز الرئيسيات عن بقية الحيوانات هي المهارة اليدوية. الطرف limb لدى الرئيسيات لم يعد مجرد أداة للحركة الانتقالية على غرار السوط flagellum أو الهدب المتحرك motile cilium أو القدم الزائفة pseudopodium، ولكنه تحول تدريجيا إلى عضو للإمساك بالأشياء وتفحصها والتأثير فيها.
هذا التغير في وظيفة الأطراف يظهر بشكل جلي لدى البشر. الطرفان العلويان لدى البشر لم يعد لهما علاقة تذكر بتحريك الجسم، أي أن وظيفتهما الأصلية تلاشت تماما.
بالنسبة للطرفين السفليين فهما ما زالا يؤديان وظيفة تحريك الجسم، ولكن الطريقة التي يؤديان بها هذه الوظيفة تغيرت كثيرا. الطريقة التي يمشي بها البشر لا تكاد تشبه الطريقة التي تمشي بها غالبية الثدييات. البشر يعتمدون المشية ثنائية القدم bipedal وليس المشية رباعية القدم quadrupedal التي تميز غالبية رباعيات الأرجل tetrapods. هناك اختلافات كبيرة بين المشيتين. مثلا البشر لا يكادون يثنون أرجلهم عند المشي، وحتى عند الوقوف أو النوم هم في الغالب لا يثنون أرجلهم. في الحقيقة البشر يبقون أرجلهم منبسطة في أغلب الأوقات. هذا يختلف عن الحال لدى الحيوانات ذات المشية رباعية الأرجل. تلك الحيوانات تبقي أرجلها مثنية في أغلب الأوقات. بما أن وظيفة الـ RNm هي ثني الأطراف فليس مستغربا أن هذه النواة فقدت تأثيرها على الطرفيين السفليين لدى البشر. تأثير هذه النواة على الطرفيين السفليين لم يعد مفيدا للبشر.
إذن النواة الحمراء كبيرة الخلايا RNm أصبحت قليلة الفائدة للبشر، لأن البشر ما عادوا يثنون أطرافهم الخلفية إلا فيما ندر. هم ما زالوا يثنون أطرافهم الأمامية، ولكنهم لا يمشون على هذه الأطراف. هذه الأطراف صارت تستخدم في الأساس للإمساك بالأشياء والتأثير فيها. وظيفة الإمساك بالأشياء والتأثير فيها تتم عبر أصابع اليدين. التحكم بأصابع اليدين يتم عبر الجهاز الدهليزي (قشرة المخ). لهذا السبب الـ RNm لم تعد عضوا مهما ولكنها صارت عضوا شبه أثري vestigial.
صفحة ويكيبيديا تقول أن النواة الحمراء تتحكم بحركة الزحف crawling لدى الأطفال الرضع، وأنها مسؤولة عن أرجحة الذراعين أثناء المشي، ولكن مصدر هذه المعلومات غير مذكور.
وظيفة النواة الحمراء صغيرة الخلايا
وظيفة النواة الحمراء صغيرة الخلايا RNp هي أقل وضوحا من وظيفة نظيرتها كبيرة الخلايا RNm. كما قلنا في الأعلى فإن اتصالات هذه النواة أوحت للباحثين أنها جزء من الجهاز الهرمي. الجهاز الهرمي هو مسؤول عن الحركات المعقدة التي تميز البشر من قبيل تحريك الأصابع والكلام. هذه الدراسة مثلا تربط بين نمو الـ RNp لدى البشر وبين تطور المهارات اليدوية والكلامية لديهم.
حسب ما فهمت فإن الـ RNp لا ترسل أية ألياف نازلة نحو الحبل الشوكي. هذه النواة هي في الأصل نواة حسية (صغيرة الخلايا parvocellular) وليست نواة تحريكية.
في الحقيقة اتصالات الـ RNp هي شبيهة باتصالات النواة الزيتونية السفلية inferior olivary nucleus، وهناك حتى اتصال مباشر بين النواتين عبر الطريق الأحمر-الزيتوني rubro-olivary tract. النواة الزيتونية السفلية (كالـ RNp) تتلقى أليافا من نواة المخيخ المسننة dentate nucleus.
بناء على التشابه في الأصل التكويني والاتصالات العصبية بين الـ RNp والنواة الزيتونية السفلية أنا أشعر بأن هاتين النواتين هما مكملتان لبعضهما ومن الممكن أنهما تؤديان نفس الوظيفة (تحديث معلومات المخيخ). النواة الزيتونية السفلية تستقبل بيانات قادمة من الحبل الشوكي، والـ RNp تستقبل بيانات قادمة من قشرة المخ. التجميع النهائي لهذه البيانات يتم في النواة الزيتونية السفلية، ومن هناك يتم إرسال إشارات التحديث أو التصحيح نحو المخيخ (والله أعلم).
في المقال القادم سأكمل الحديث في موضوع الحركة الانتقالية بواسطة الأطراف.
الدماغ البشري (10)
في المقال الأخير من هذه السلسلة بدأنا الحديث عن مقطع الدماغ الأوسط (midbrain) mesencephalon وتحدثنا عن ساقي المخ crura cerebri وبدأنا الحديث عن الغطاء tegmentum.
في ذلك المقال تحدثنا عن “المنطقة الغطائية المركزية” central tegmental area التي تضم التشكل الشبكي reticular formation، وبينا أن بعض أنوية التشكل الشبكي هي في الحقيقة متسربة إلى خارج هذه المنطقة.
أنوية الرفاء raphe nuclei هي من أنوية التشكل الشبكي ولكنها تقع على الجهة الإنسية medial للطرق العصبية الصاعدة التي تحد المنطقة الغطائية المركزية من الجهة البطنية (مثل الطريق المسنّن-الثلمي dentatothalamic tract والشريط الإنسي medial lemniscus).
النواة بين السويقية
على الجهة البطنية لأنوية الرفاء توجد نواة تسمى “النواة بين السويقية” interpeduncular nucleus. هذه النواة هي مكونة بمعظمها من خلايا تثبيطية مفرزة للـ GABA. هذه النواة هي مرتبطة وظيفيا بجزء من الدماغ البيني diencephalon يسمى “العُنَيِّن” habenula (العُنَيِّن يقع على الجهة الإنسية الظهرية للثلم thalamus بالقرب من الغدة الصنوبرية pineal gland).
الدراسات بينت أن العنين يلعب دورا هاما في تنظيم السلوك والتفكير. سوف نؤجل الحديث عن وظيفة العنين إلى وقت لاحق، ولكن الآن سنقول باختصار ما يلي: منظومة العنين هي مثبطة للسلوك والتفكير، أي أنها تؤدي دورا معاكسا لدور المنظومات المفرزة للنواقل أحادية الأمين monoamines والـ acetylcholine (النواقل أحادية الأمين تشمل الـ norepinephrine والـ dopamine والـ serotonin).
النواة بين السويقية تتلقى أليافا نازلة من نواة “العنين الإنسيmedial habenula “. بعض الباحثين أتلفوا نواة العنين الإنسي لدى فئران المختبر، والنتيجة كانت أن هذه الفئران أصبحت ذات نشاط زائد hyperactive، ومندفعة impulsive/compulsive (بسبب النشاط الزائد لمنظومة الـ dopamine)، وزائدة القلق anxiety (بسبب النشاط الزائد لمنظومة norepinephrine)، وأصبحت تتخذ قرارات متسرعة وغير حكيمة delay discounting (بسبب النشاط الزائد لمنظومة الـ norepinephrine)، كما أنها عانت من مشاكل في عمل الذاكرة طويلة الأجل long-term memory (ربما بسبب الخلل في عمل منظومة الـ serotonin). هذه النتائج تدل على وظيفة النواة بين السويقية—هذه النواة هي مثبطة للأنظمة التنشيطية الموجودة في الدماغ الأوسط.
المادة السوداء
على الجهة الوحشية lateral للنواة بين السويقية هناك نواة عريضة تسمى “المادة السوداء” substantia nigra. هذه النواة هي في الحقيقة نواتان اثنتان وليست نواة واحدة؛ القسم البطني من المادة السوداء يسمى “القسم الشبكي” pars reticulata وهو مكون من خلايا تثبيطية مفرزة للـ GABA. القسم الظهري من المادة السوداء يسمى “القسم المضغوط” pars compacta وهو مكون من خلايا مفرزة للـ dopamine.
وظيفة القسمين الشبكي والمضغوط هي متناقضة. القسم المضغوط هو أحد مواقع المنظومة الدوبامينية (الطرق المفرزة للدوبامين dopaminergic pathways) المسؤولة عن توليد الرغبة أو الشهية أو الإرادة أو الحاجة إلخ (كل هذه الألفاظ استخدمت في وصف عمل الطرق الدوبامينية). القسم المضغوط من المادة السوداء ينتمي لمنظومة دماغية تسمى “العقد القاعدية” basal ganglia. هذه المنظومة تضم عددا من الأنوية والعقد المسؤولة عن اختيار السلوك المناسب للإنسان أو الحيوان بناء على تجاربه السابقة. القسم المضغوط يلعب في هذه المنظومة دورا محفزا للسلوك.
القسم الشبكي من المادة السوداء ينتمي أيضا لمنظومة العقد القاعدية، ولكنه يلعب في هذه المنظومة دورا مثبطا للسلوك (إلى جانب أنوية تثبيطية أخرى من قبيل القسم الداخلي من الكرة الشاحبة internal globus pallidus والنواة تحت الثلمية subthalamic nucleus).
إذن وظيفة القسم الشبكي هي معاكسة لوظيفة القسم المضغوط؛ وظيفة القسم الشبكي تشبه وظيفة النواة بين السويقية التي أشرنا إليها في الأعلى: كلتا هاتين النواتين هما تثبيطيتان مفرزتان للـ GABA، ولكن الفرق بينهما هو أن القسم الشبكي من المادة السوداء يثبط السلوك ولا يؤثر كثيرا على التفكير (حسب علمي). أيضا القسم المضغوط من المادة السوداء هو محفز للسلوك ولا يؤثر كثيرا على التفكير (حسب علمي).
في المستقبل سوف أتحدث عن عمل هذه المنظومات بتوضيح أكثر.
المنطقة الغطائية البطنية
المقطع التالي هو للدماغ الأوسط عند مستوى التليلتين العلويتين superior colliculi:
هذا المقطع لا يختلف جوهريا عن مقطع الدماغ الأوسط الذيلي سوى في أمرين:
- ظهور النواة الحمراء red nucleus في القسم البطني من الغطاء
- ظهور المنطقة الغطائية البطنية ventral tegmental area
بالنسبة للجسم الركبي الإنسي medial geniculate body فهو يقع في مؤخرة الثلم thalamus وليس جزءا من الدماغ الأوسط (رغم ظهوره في هذا المقطع).
المنطقة المسماة “المنطقة الغطائية البطنية” ventral tegmental area (اختصارا VTA) تحوي نسبة عالية (50-60%) من الخلايا المفرزة للـ dopamine، وتحوي أيضا خلايا مفرزة للـ GABA، أي أن تكوينها يشبه تكوين المادة السوداء، وهي في الحقيقة تشترك في الأصل الجنيني مع المادة السوداء. منطقة VTA هي إحدى أهم مواقع المنظومة الدوبامينية في الدماغ البشري. الخلايا المفرزة للـ dopamine في VTA هي محفزة للتفكير، ودورها تحديدا هو تحفيز الرغبة أو الشهية أو الإرادة أو الحاجة إلخ.
الألياف المفرزة للـ dopamine الصادرة من القسم المضغوط من المادة السوداء تذهب نحو “المُخطَّط” striatum، الذي هو جزء من العقد القاعدية basal ganglia يقع في قلب نصف الكرة المخية، وأما الألياف المفرزة للـ dopamine الصادرة من VTA فهي تذهب إلى بقية مناطق الدماغ الأمامي (forebrain) prosencephalon.
الألياف المفرزة للـ dopamine الصادرة من VTA توصف بأنها محفزة للرغبة والشهية ونحو ذلك، ولكن بناء على نتائج الدراسات الحديثة أنا أظن أن الوظيفة الدقيقة لهذه الألياف هي تحفيز الخيال والتفكير القائم على الخيال.
الدراسات الحديثة بينت أن الألياف المفرزة للـ dopamine الصادرة من VTA هي على ما يبدو مسؤولة عن الخيال بكل أشكاله. كلمة “الخيال” بمعناها الواسع يمكن أن تشمل الهلوسات، وأيضا الأحلام التي يراها الإنسان أثناء نومه. الألياف المفرزة للـ dopamine المسؤولة عن تحفيز الهلوسات والأحلام تسمى “الطريق الأوسط-الحَوْفي” mesolimbic pathway. النشاط الزائد في هذا الطريق هو ربما سبب الهلوسات التي يراها المصابون بمرض schizophrenia.
عبارة “التفكير القائم على الخيال” تشمل نوعين من التفكير على الأقل:
- التفكير الرغبوي (تخيل الإنسان للأشياء التي يريدها)
- التفكير الإبداعي (تخيل الإنسان لكيفية الوصول إلى الأشياء التي يريدها)
هذان النوعان من التفكير يحصلان في القشرة أمام الجبهية prefrontal cortex، التي هي مقر “ذاكرة العمل” working memory. الألياف المفرزة للـ dopamine التي تغذي القشرة أمام الجبهية تسمى “الطريق الأوسط-القشري” mesocortical pathway.

النظريات الحديثة حول مرض schizophrenia (السكيزوفرينيا أو الشيزوفرينيا) ترى أن من أسباب هذا المرض انقطاع التواصل العصبي بين المناطق المخّية المولدة للخيال (التي يغذيها الطريق الأوسط-الحَوْفي mesolimbic pathway) والمناطق المخّية المسؤولة عن معالجة الخيال (التي يغذيها الطريق الأوسط-القشري mesocortical pathway). انقطاع التواصل بين هاتين المنظومتين (أو بالأحرى وجود خلل في هذا التواصل) يجعل الإنسان عاجزا عن إدراك حقيقة خيالاته: هو يتصور أنها مدركات حسية نابعة من العالم الخارجي ولا يدري أنها خيالات نابعة من مناطق أخرى في مخه.
ما سبق هو تلخيص بسيط يدل على أهمية الخلايا المفرزة للـ dopamine في التفكير البشري. في مقالات قادمة سوف أتحدث بمزيد من التفصيل عن هذه القضايا الهامة.
أصل الخلايا المفرزة للـ dopamine
في الأعلى استعرضنا أهم الأنوية المفرزة للـ dopamine في الدماغ الأوسط (القسم المضغوط pars compacta من المادة السوداء substantia nigra، والمنطقة الغطائية البطنية ventral tegmental area)، واستعرضنا بعض الأنوية المثبطة لهذه الأنوية (القسم الشبكي pars reticulata من المادة السوداء الذي يعاكس القسم المضغوط، والنواة بين السويقية interpeduncular nucleus التي تعاكس المنطقة الغطائية البطنية وأنوية الرفاء raphe nuclei وغير ذلك).
ذكرنا في الأعلى أن المادة السوداء والمنطقة الغطائية البطنية لهما أصل جنيني واحد، بمعنى أنهما في المرحلة الجنينية تَكُونان نواة واحدة وبعد ذلك هذه النواة تنقسم إلى نواتين. أنا أظن أن أصل هذه النواة يعود إلى القسم الإنسي medial من التشكل الشبكي، ربما في نفس موقع النواة بين السويقية interpeduncular nucleus.
النواة بين السويقية ربما تعود إلى نفس هذا الأصل المشترك. المادة السوداء تحوي خلايا مفرزة للـ GABA في قسمها الشبكي، والمنطقة الغطائية البطنية تحوي أيضا خلايا مفرزة للـ GABA كما ذكرنا في الأعلى. من الممكن أن نتخيل أن كل هذه الأنوية تتحدر من أصل واحد كان يحوي خلايا مفرزة للـ dopamine وأخرى مفرزة للـ GABA. هذا الأصل المشترك كان يقع ربما على الجهة البطنية لأنوية الرفاء.
أشرت في مقال سابق إلى الدراسة التي بحثت تواجد الخلايا المفرزة للـ serotonin في الحيوانات ثنائية الجانب bilaterians. حسب تلك الدراسة فإن الخلايا المفرزة للـ serotonin كانت توجد في وسط الجهاز العصبي لثنائيات الجانب الأولى urbilaterians.

حسب هذه الدراسة فإن جميع أنظمة الناقلات أحادية الأمينmonoamine (التي تشمل الـ norepinephrine والـ dopamine والـ serotonin) هي متحدرة من أصل واحد:
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3070214/
Many of the molecular components of DA [dopamine] systems, such as biosynthetic enzymes, transporters, and receptors, are shared with those of other monoamine systems, suggesting the common origin of these systems.
هذه الدراسة بحثت تحديدا في تطور أنظمة الـ dopamine في الحيوانات الحبلية chordates. الخلايا المفرزة للـ dopamine لدى الحيوانات الحبلية توجد في أربعة مواقع:
- البصلة الشمية olfactory bulb
- شبكية العين retina
- الدماغ البيني diencephalon (لدى الثدييات في “ما تحت الثلم” hypothalamus)
- الدماغ الأوسط mesencephalon
الباحثون يقسمون الخلايا المفرزة للـ dopamine في الدماغ إلى مجموعات مرقمة. الرسم التالي يظهر هذه المجموعات في الدماغ البشري:

حسب الدراسة المشار إليها فإن أصل الخلايا المفرزة للـ dopamine يعود إلى الدماغ الأمامي prosencephalon. مؤلفا الدراسة يريان أن الخلايا المفرزة للـ dopamine كانت لدى الحبليات الباكرة موجودة حول القسم الأمامي من تجويف الأنبوب العصبي ضمن منطقة مكونة من نوعين من الخلايا:
- خلايا عصبية-هرمونية neuroendocrine cells (خلايا عصبية تفرز الهرمونات في الدم)
- خلايا مستقبلة للضوء photoreceptor cells
الرسم التالي (مصدره نفس الدراسة) يوضح المنطقة التي نتحدث عنها (في الكائن الموجود على جهة اليسار):

الكائن (البدائي) الموجود على جهة اليسار يحوي في دماغه الأمامي منطقة مكونة من نوعين متداخلين من الخلايا: خلايا عصبية-هرمونية تظهر باللون الأحمر، وخلايا مستقبلة للضوء تظهر باللون الأزرق. حسب الدراسة فإن بعض الخلايا العصبية-الهرمونية كانت مفرزة للـ dopamine وبعضها الآخر كانت مفرزة لهرمونات أخرى (من قبيل الهرمون الحيواني المسمى vasotocin). وظيفة الخلايا العصبية-الهرمونية كانت إفراز الهرمونات بناء على الإشارات الواردة من الخلايا المستقبلة للضوء المتداخلة معها.
إذن الدراسة تعتبر أن الخلايا المفرزة للـ dopamine هي من الخلايا العصبية-الهرمونية neuroendocrine cells. أنا أشك في صحة هذا التصنيف، لأن الخلايا المفرزة للـ dopamine الموجودة في الدماغ البشري (والحيواني أيضا على الأغلب) لا تطرح الـ dopamine في الدم. الخلايا المفرزة للـ dopamine الموجودة في ما تحت الثلم hypothalamus تتحكم بإفراز الهرمونات من الخلايا العصبية-الهرمونية (الحقيقية)، ولكنها لا تطرح أية هرمونات في الدم بشكل مباشر. إذن هذه الخلايا هي مجرد خلايا عصبية ربطية وليست خلايا عصبية-هرمونية.
الملفت هو أن مؤلفي الدراسة لم يذكرا شيئا حول الخلايا المستقبلة لحس الشم. نفس الأمر لاحظته في الدراسة المتعلقة بثنائيات الجانب التي أشرت لها في مقال سابق. كلتا الدراستين أشارتا إلى وجود مستقبلات ضوئية في الدماغ الأمامي للحبليات الباكرة وثنائيات الجانب الباكرة، ولكنهما لم تذكرا شيئا عن المستقبلات الشمية.
حسب هذه الدراسة فإن جينات المستقبلات الشمية (OR) olfactory receptor هي قديمة وتعود إلى زمن الحبليات الأولى. أنا أشرت في مقال سابق إلى حيوان حبلي بدائي التكوين يسمى lancelet أو amphioxus، وقلت وقتها أن هذا الحيوان هو ربما الأكثر شبها بالحيوانات الحبلية الأولى. حسب الدراسة المشار إليها فإن جينات المستقبلات الشمية OR هي موجودة لدى هذا الحيوان:
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2817399/
Amphioxus is called an “acraniate,” meaning a headless organism. It lacks an identifiable olfactory organ, and almost nothing is known of its sensitivity to chemical stimuli (Lacalli 2004). Nevertheless, many vertebrate-type OR-like genes were found in the amphioxus genome. Therefore, the origin of vertebrate-type OR genes can be traced back to the common ancestor of chordates. (Recently, Grus and Zhang [2009] suggested the same time for the origin of the OR gene family.) Satoh (2005) reported that at least one OR-like gene is broadly expressed in bipolar neurons embedded within the rostral epithelium of adult amphioxus. Further studies will be necessary to examine the cell types in which amphioxus OR genes are expressed.
بناء على هذا الكلام فإن حيوان الـ lancelet يملك جينات المستقبلات الشمية، ومؤخرا عُثر في هذا الحيوان على مستقبلات شمية مطمورة بين خلايا جلده المنقاري rostral epithelium. إذن المستقبلات الشمية كانت موجودة لدى الحبليات الباكرة وكانت تقع في الجهة المنقارية.
السؤال هو ما العلاقة بين المستقبلات الشمية وبين الخلايا العصبية-الهرمونية والخلايا المفرزة للـ dopamine التي كانت تقع في الدماغ الأمامي؟ الدراسات لم تذكر شيئا حول هذا الموضوع، ولكن على الأغلب أن هناك علاقة. لو نظرنا إلى عالم الحيوانات الفقارية فسنجد ظاهرة فيزيولوجية مهمة هي الفرمونات pheromones. الفرمونات هي جزيئات كيميائية شبيهة بالهرمونات ولكنها تفرز في الهواء وتنتقل من حيوان إلى حيوان آخر. الحيوان يستقبل الفرمونات عبر المستقبلات الشمية. استقبال الفرمونات يولد في الدماغ إشارات عصبية تؤثر في السلوك “الغريزي” المرتبط بمنطقة ما تحت الثلم hypothalamus (كالسلوك الجنسي مثلا)، وبالتالي الفرمونات هي وسيلة للتواصل بين الحيوانات المختلفة التي تنتمي إلى نفس النوع species.
الدراسة المتعلقة بتطور أنظمة الـ dopamine التي أشرنا إليها في الأعلى تجاهلت المستقبلات الشمية. حسب تلك الدراسة فإن منطقة الخلايا العصبية-الهرمونية والخلايا المفرزة للـ dopamine في الدماغ الأمامي كانت “منطقة استقبال ضوئي”photoreceptive territory :
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3070214/#s4
[…] DA cells in the ancestral chordate were located in a photoreceptive territory of the anterior neural tube, which is homologous to both retina and hypothalamic region […] These DA cells were probably multifunctional and able to modulate neural functions such as sensory input, motor control, and hormonal regulation.
[…]
Recent data about the DA systems of ascidia suggest that DA cells of the sensory vesicle on the one hand, and DA cells of the hypothalamus and the retina of vertebrates on the other hand, may have originated from a common neuroendocrine, photoreceptive territory surrounding the ventricle of the neural tube in ancestral chordates.
هذه الدراسة تقول أن منطقة الخلايا العصبية-الهرمونية والخلايا المفرزة للـ dopamine (“منطقة الاستقبال الضوئي”) تمايزت لاحقا وأنتجت ما تحت الثلم hypothalamus والحويصلات البصرية (وهذه بدورها أنتجت شبكيتي العينين). الدراسة لا تذكر شيئا عن أصل البصلتين الشميتين olfactory bulbs، ولكن الملفت هو أن الدراسة تعيد أصل الخلايا المفرزة للـ dopamine الموجودة في البصلتين الشميتين إلى نفس منطقة الخلايا العصبية-الهرمونية والخلايا المفرزة للـ dopamine التي كانت تقع حول القسم الأمامي للأنبوب العصبي:
Neural precursors of the DA cells of the olfactory bulb arise from the subventricular zone of the telencephalon and migrate to the olfactory bulb by the rostral migratory stream. This neurogenesis persists all life long in all vertebrates studied so far, and it is strongly affected by activity (Lazarini and Lledo, 2011).
منطقة الخلايا العصبية-الهرمونية والخلايا المفرزة للـ dopamine في الدماغ الأمامي للحبليات الباكرة كانت على ما أظن تتعامل مع حس الشم إلى جانب حس البصر، ولكن من الممكن أن حس الشم كان هامشيا مقارنة مع حس البصر. الدراسة المتعلقة بحس الشم التي أشرنا إليها في الأعلى ذكرت أن حس الشم لدى حيوان lancelet هو هامشي أو غير واضح. إذن يمكننا أن نتخيل أن حس الشم لدى الحبليات الباكرة كان هامشيا مقارنة مع حس البصر، ولكنه كان موجودا.
منطقة الخلايا العصبية-الهرمونية والخلايا المفرزة للـ dopamine في الدماغ الأمامي تمايزت لاحقا وأنتجت الأعضاء الحسية التالية:
- البصلتان الشميتان olfactory bulbs (في الدماغ النهائي telencephalon)
- الحويصلان البصريان optic vesicles (ولاحقا الشبكيتان retinae) (في الدماغ البيني diencephalon)
هذه الأعضاء هي ليست كل الأعضاء الحسية التي نشأت من الدماغ الأمامي؛ هناك أعضاء أخرى أقل أهمية أو أقل شهرة. من هذه الأعضاء “العين الثالثة” أو “العين الجدارية” التي توجد لدى بعض الحيوانات الفقارية (والتي أشرت إليها في مقال سابق). الحيوانات الفقارية تملك أيضا بصلتين شميتين إضافيتين تسميان “البصلتين الشميتين الملحقتين” accessory olfactory bulbs. هاتان البصلتان تقعان فوق البصلتين الشميتين الرئيسيتين وهما تتصلان مع مستقبلات حسية خاصة توجد ضمن عضوين أنفيين يسميان (VNO) vomeronasal organ.


لو أردنا الحديث بشكل دقيق فإن الأعضاء الحسية التي نشأت من الدماغ الأمامي هي ما يلي:
- 4 بصلات شمية من الدماغ النهائي (بصلتان أساسيتان وبصلتان ملحقتان)
- 3 حويصلات بصرية من الدماغ البيني (العينان الجانبيتان والعين الجدارية)
هذه الأعضاء الحسية تحوي خلايا عصبية وعصبية-هرمونية مشتقة من خلايا المنطقة البدائية التي كانت تقع حول مقدمة تجويف الأنبوب العصبي. الخلايا التي بقيت حول مقدمة تجويف الأنبوب العصبي أنتجت لدى الفقاريات المتأخرة ما يسمى بالدماغ البيني diencephalon، وخاصة الجزء المسمى “ما تحت الثلم” hypothalamus. هناك قسم من هذه الخلايا تمدد أيضا نحو الدماغ الأوسط وأنتج الخلايا المفرزة للـ dopamine الموجودة في الدماغ الأوسط.
مما سبق يتبين أن أصل الخلايا المفرزة للـ dopamine يعود ربما إلى منطقة الخلايا العصبية-الهرمونية والخلايا المفرزة للـ dopamine الموجودة في الدماغ الأمامي (لدى الثدييات هي موجودة في ما تحت الثلم hypothalamus).
لو نظرنا إلى الرسم التالي فسنرى أن الخلايا المفرزة للـ dopamine الموجودة في الدماغ الأمامي هي استمرار منقاري لأنوية الرفاء:

أنوية الرفاء (الخلايا المفرزة للـ serotonin) تظهر على هذا الرسم باللون الأحمر. على الجهة المنقارية لهذه الخلايا توجد الخلايا العصبية-الهرمونية neuroendocrine التي تظهر باللون البرتقالي (هذه الخلايا توجد لدى الثدييات في ما تحت الثلم hypothalamus). بناء على كلام الدراسة التي أشرنا إليها في الأعلى فإن الخلايا المفرزة للـ dopamine في الدماغ الأمامي هي متداخلة مع الخلايا العصبية-الهرمونية، أي أنها جزء من الخلايا ذات اللون البرتقالي. الخلايا ذات اللون الأخضر هي خلايا “الساعة اليومية” circadian clock (توجد لدى الثدييات في النواتين فوق التصالبيتين suprachiasmatic nuclei في القسم المنقاري من ما تحت الثلم hypothalamus). الخلايا ذات اللونين الأبيض والأصفر هي مستقبلات ضوئية.
بما أن الخلايا المفرزة للـ dopamine هي على ما يبدو مجرد امتداد منقاري (في الدماغ الأمامي) للخلايا المفرزة للـ serotonin فهذا يطرح احتمالية الأصل المشترك لهذه الخلايا. هذه الاحتمالية هي واردة كما قالت الدراسة التي نقلت منها في الأعلى. الـ dopamine والـ serotonin هما جزيئان متشابهان كيميائيا: كلاهما ينتميان إلى أحاديات الأمين monoamines. الـ dopamine هو مشتق كيميائيا من حمض أميني يسمى tyrosine، والـ serotonin هو مشتق من حمض أميني يسمى tryptophan.

أصل الخلايا المفرزة للـ norepinephrine
في الدماغ الأوسط توجد خلايا مفرزة لناقل آخر من أحاديات الأمين هو الـ norepinephrine. لو نظرنا في المقطعين C و D من الشكل التالي فسنرى أن الموضع الأزرق locus coeruleus (المفرز للـ norepinephrine) يقع على الجهة الظهرية لأنوية الرفاء:
أنا بصراحة لم أقرأ أي شيء مفيد حول أصل الخلايا المفرزة للـ norepinephrine، ولذلك ما سأقوله الآن هو ليس أكثر من اجتهادات أو تخرصات شخصية.
الـ norepinephrine هو من الناحية الكيميائية شديد الشبه بالـ dopamine. كلا هذين الجزيئين ينتميان إلى تصنيف كيميائي يسمى catecholamines (هذا التصنيف يضم أيضا الـ epinephrine الذي هو مشتق كيميائيا من الـ norepinephrine). الـ dopamine هو مصدر الـ norepinephrine في الجسم البشري. تحويل الـ dopamine إلى norepinephrine يتم عبر تفاعل كيميائي بسيط ينفذه أنزيم واحد كما يظهر في الرسم التالي:

هذا الرسم يوحي بوجود صلة قرابة وثيقة بين الـ dopamine والـ norepinephrine. من الممكن أن هذين الناقلين يتحدران من أصل مشترك حديث نسبيا.
حسب ما فهمت من بعض المصادر فإن الـ norepinephrine هو غير موجود لدى الحيوانات غير الفقارية. الحيوانات غير الفقارية تملك ناقلا آخر يسمى (OA) octopamine. هذا الناقل يؤدي نفس الوظائف التي يؤديها الـ norepinephrine لدى الحيوانات الفقارية. بعض الباحثين حقنوا حيوان الـ lobster بالـ serotonin، فكانت النتيجة أن الحيوان صار يتصرف وكأنه الزعيم في مجتمعه، ولكنهم عندما حقنوه بالـ octopamine تغير سلوكه وصار يتصرف وكأنه فرد حقير في مجتمعه. إذن من الممكن أن الـ octopamine هو مولد للخوف أو القلق، وهي نفس وظيفة الـ norepinephrine لدى الفقاريات.
الـ octopamine هو جزيء أحادي الأمين مشتق من الـ tyrosine، ولكنه ليس مشتقا من الـ dopamine.

على ما يبدو فإن ظهور الفقاريات (أو ربما الحبليات) ترافق مع التخلي عن الـ octopamine والانتقال نحو الـ norepinephrine، ولكن السؤال هو ما سبب هذا التغيير؟
أنا بصراحة لا أعرف ما هو السبب، ولكن إحدى الأفكار التي طُرحت هي أن الانتقال نحو الـ norepinephrine هدفه الربط بين إنتاج الـ norepinephrine وإنتاج الـ dopamine.
إنتاج الـ octopamine في الجسم يتم بمعزل عن إنتاج الـ dopamine، وبالتالي من الممكن نظريا أن يكون هناك حيوان يملك في جهازه العصبي كمية كبيرة من الـ octopamine وكمية منخفضة من الـ dopamine. في المقابل إنتاج الـ norepinephrine هو مرتبط بإنتاج الـ dopamine، وبالتالي لا يمكن أن يكون هناك حيوان يملك في جهازه العصبي كمية كبيرة من الـ norepinephrine وكمية منخفضة من الـ dopamine، لأن إنتاج الـ norepinephrine يمر عبر إنتاج الـ dopamine.
أنا ذكرت سابقا أن الـ norepinephrine هو مسؤول عن اتخاذ القرارات، وأما الـ dopamine فهو مسؤول عن تشغيل منظومة العقد القاعدية basal ganglia التي تنظم السلوك بناء على التجارب السابقة التي مر بها الحيوان. الحيوان الذي يعتمد على منظومة العقد القاعدية في اتخاذ قراراته هو أكثر أمانا من غيره، لأنه يعتمد على خبراته وتجاربه المتراكمة (مثلا هو يعلم ما هي الأشياء الخطيرة حتى يتجنبها وما هي الأشياء الجيدة حتى يطلبها). الحيوان الذي لا يستخدم منظومة العقد القاعدية هو عمليا يتصرف وكأنه حشرة، بمعنى أنه يتحرك في بيئته دون أية معرفة مسبقة بالأشياء الخطيرة والأشياء غير الخطيرة. هذا الحيوان هو معرض للأذى أكثر من الحيوان الذي يستفيد من تجاربه السابقة.
لو فرضنا أن هناك حيوانا فقاريا لديه نسبة عالية من الـ octopamine ونسبة منخفضة من الـ dopamine فما هو السلوك المتوقع لهذا الحيوان؟ هذا الحيوان سوف يتخذ قراراته بسرعة زائدة ودون اعتماد كبير على منظومة العقد القاعدية، أي أنه سوف يكون حيوانا متسرعا ومتهورا، وبالتالي مهددا بالفناء.
ربما يكون هذا هو السبب الذي أدى لانقراض الحيوانات الفقارية المعتمدة على الـ octopamine. هذه الحيوانات انقرضت لأنها كانت تتصرف على نحو متهور ودون استفادة من تجاربها السابقة. وجود نسبة عالية من الـ octopamine ونسبة منخفضة من الـ dopamine يقلل مناسبة fitness الحيوان (وفق مفاهيم نظرية التطور evolution فإن الحيوان غير المناسب unfit هو معرض للانقراض أكثر من الحيوان المناسب fit).
بناء على هذه النظرية (التي هي نظرية شخصية من عندي ولم أقرأها في أي مكان) فإن التخلى عن الـ octopamine ترافق مع ظهور منظومة العقد القاعدية basal ganglia لدى الحيوانات الفقارية الباكرة. الحيوانات الفقارية التي استفادت من منظومة العقد القاعدية هي التي بقيت وتكاثرت، وأما الحيوانات الفقارية التي لم تستفد من هذا الاختراع الجديد وظلت تتصرف كما لو أنها حيوانات غير فقارية فكان مصيرها الانقراض والفناء.
الاستفادة من منظومة العقد القاعدية تعني حكما الاعتماد الكبير على الـ dopamine، لأن هذا الناقل هو الذي يشغّل منظومة العقد القاعدية. الفقاريات الموجودة حاليا لا تتخذ أي قرار دون مشاورة العقد القاعدية، ولهذا السبب هذه الحيوانات تعتمد على الـ norepinephrine (المشتق من الـ dopamine) ولا تعتمد على الـ octopamine (المستقل عن الـ dopamine). الاعتماد الكامل على الـ norepinephrine في اتخاذ القرارات يعني أن تجاهل العقد القاعدية هو أمر غير ممكن، لأن الوصول إلى اتخاذ قرار عبر إفراز الـ norepinephrine يتطلب حكما إفراز الـ dopamine، وبالتالي لا يمكن لقرار أن يتخذ دون إفراز الـ dopamine.
مما سبق نستنتج أن الـ norepinephrine هو ناقل حديث نسبيا، وهو مشتق من الـ dopamine، بالتالي من الممكن أن الخلايا المفرزة للـ norepinephrine (الموضع الأزرق) كانت في الأصل مفرزة للـ dopamine. هذا يوصلنا إلى أن أصل الموضع الأزرق ربما يعود إلى الخلايا المفرزة للـ dopamine الموجودة في الدماغ الأمامي. طبعا هذا الكلام هو مجرد تخيلات من عندي وليس مبنيا على كلام قرأته في مصادر علمية.
أنا لا أعرف ما هي الخلايا المفرزة للـ octopamine لدى الحيوانات غير الفقارية، ولكن الـ octopamine هو من أحاديات الأمين، وبالتالي المفترض هو أنه يشترك في الأصل مع الـ serotonin والـ dopamine.
في مقال سابق أشرت إلى دراسة تحدثت عن تطور الجهاز العصبي ذاتي الحكم autonomic nervous system لدى الفقاريات. حسب ما ورد في تلك الدراسة فإن الفقاريات البدائية لم تكن تملك عقدا محيطية مفرزة للـ norepinephrine. جميع الأعصاب الحشوية الصادرة من الحبل الشوكي لدى تلك الحيوانات كانت مفرزة للـ acetylcholine. الأعصاب المتعاطفة sympathetic المفرزة للـ norepinephrine كانت تخرج من جذع الدماغ (ربما كانت تخرج من “الحقل الغطائي الجانبي” lateral tegmental field الذي هو حسب علمي المنطقة الوحيدة المفرزة للـ norepinephrine في جذع الدماغ إلى جانب الموضع الأزرق).
إذن وجود العقد المحيطية المفرزة للـ norepinephrine هو ظاهرة حديثة نسبيا.
أحاديات أمين أخرى
في الأعلى أنا تحدثت عن ناقلين مهمين من أحاديات الأمين هما الـ dopamine والـ norepinephrine. بناء على ما ورد في الأعلى فإن أصل الخلايا المفرزة لهذين الناقلين ربما يعود إلى الخلايا المفرزة للـ dopamine الموجودة في الدماغ الأمامي prosencephalon.
في الدماغ البشري توجد نواقل أخرى من أحاديات الأمين؛ من هذه النواقل مثلا الناقل المسمى melatonin. هذا الناقل يفرز من خلايا الغدة الصنوبرية pineal gland الموجودة في مؤخرة الثلم thalamus والتي تشكل جزءا من منظومة تسمى “ما فوق الثلم” epithalamus. منظومة “ما فوق الثلم” هي مرتبطة تطوريا بالعين الثالثة أو العين الجدارية parietal eye التي أشرنا إليها آنفا.
الـ melatonin من الناحية الكيميائية هو وثيق الصلة بالـ serotonin. العلاقة بين الـ serotonin والـ melatonin تشبه العلاقة بين الـ dopamine والـ norepinephrine.

وجود خلايا مفرزة للـ melatonin ضمن الغدة الصنوبرية (وثيقة الصلة بالعين الجدارية) يتوافق مع النظرية التي ذكرناها في الأعلى حول الدور الأصلي للخلايا المفرزة لأحاديات الأمين في الدماغ الأمامي: هذه الخلايا كانت في الأصل مسؤولة عن التعامل مع المستقبلات الضوئية (وربما أيضا مع المستقبلات الشمية).
على ما أظن فإن وجود الـ melatonin ضمن خلايا الدماغ الأمامي المفرزة لأحاديات الأمين يدعم النظرية التي تقول بوجود أصل مشترك لجميع الخلايا المفرزة لأحاديات الأمين. الـ melatonin هو وثيق الصلة بالـ serotonin؛ وجود الخلايا المفرزة للـ melatonin ضمن الخلايا المفرزة لأحاديات الأمين في الدماغ الأمامي يوحي بأن بعض هذه الخلايا ربما كانت في الماضي مفرزة للـ serotonin.
في القسم الظهري من ما تحت الثلم hypothalamus (تحديدا في النواة الحدبية-الحلمية tuberomammillary nucleus) توجد خلايا مفرزة لناقل أميني يسمى histamine. هذا الناقل هو شبيه بأحاديات الأمين ولكنه لا ينتمي إليها كيميائيا. هذا الناقل يلعب دورا كبيرا خارج الجهاز العصبي (هو مهم خاصة في الجهاز المناعي، وهو يلعب دورا مهما في الاستجابة المناعية التي تسمى شعبيا “الحساسية” allergy)، ولكن وجود الـ histamine في الجهاز العصبي المركزي ينحصر في النواة الحدبية-الحلمية.
الوظيفة الأساسية للـ histamine المفرز من النواة الحدبية-الحلمية هي التنشيط activation أو تحفيز اليقظة (لهذا السبب فإن “مضادات الهيستامين” antihistamines المستخدمة في علاج “الحساسية” تسبب النعاس)، وعلى ما يبدو فإن إفراز الـ histamine من النواة الحدبية-الحلمية هو مرتبط بالإيقاع اليومي circadian rhythm، أي أنه مرتبط بشدة الضوء الساقط على العينين. الـ histamine المفرز من النواة الحدبية-الحلمية يساهم في تنظيم إفراز بعض الهرمونات من الخلايا العصبية-الهرمونية الموجودة في ما تحت الثلم، ويساهم في تنظيم بعض المنعكسات الحشوية المعقدة التي تدار من ما تحت الثلم.
وظائف الـ histamine المفرز من النواة الحدبية-الحلمية هي الوظائف الكلاسيكية لأحاديات الأمين المفرزة من خلايا الدماغ الأمامي؛ أحاديات الأمين المفرزة من خلايا الدماغ الأمامي هي عموما منشطة أو محفزة لليقظة، وإفرازها كان في الأصل مرتبطا بشدة الضوء الساقط على المستقبلات الضوئية، وهي تساهم في تنظيم إفراز الهرمونات من الخلايا العصبية-الهرمونية الموجودة في ما تحت الثلم، وتساهم في تنظيم بعض المنعكسات الحشوية المعقدة التي تدار من ما تحت الثلم. هذه الوظائف هي الوظائف الأصلية للـ dopamine قبل أن تتمدد بعض الخلايا المفرزة له نحو الدماغ الأوسط وتكتسب وظائف جديدة من قبيل تحفيز السلوك الجيد وتحفيز التفكير الخيالي (الـ dopamine المفرز من خلايا ما تحت الثلم hypothalamus عند الإنسان هو مثبط لإفراز هرمون prolactin المسؤول عن تحفيز إفراز الحليب من الثديين، ويبدو أيضا أنه مثبط لإفراز هرمونGnRH المسؤول عن تحفيز إنتاج خلايا التكاثر الجنسي في المبيضين والخصيتين، أي أن الـ dopamine المفرز في ما تحت الثلم هو على ما يبدو مثبط للتكاثر الجنسي. هذه الوظيفة تشبه وظيفة أحاديات الأمين الأخرى: الـ serotonin والـ norepinephrine كلاهما مثبطان للرغبة الجنسية، والـ norepinephrine هو مسؤول عن الذروة الجنسية orgasm، أي أنه ينهي العملية الجنسية).
الـ histamine هو جزيء “شقيق” لأحاديات الأمين، بمعنى أنه يشبه أحاديات الأمين كيميائيا وإن كان لا ينتمي إليها، وهو يؤدي نفس الوظائف الكلاسيكية لأحاديات الأمين المفرزة في ما تحت الثلم. على ما يبدو فإن تطور هذا الناقل هو نفس تطور أحاديات الأمين المفرزة في الدماغ الأمامي: هو في الأصل كان مسؤولا عن التعامل مع مستقبل ضوئي.
الـmelatonin هو استثنائي بين أحاديات الأمين المفرزة في الدماغ الأمامي من حيث أن إفرازه يزداد في غياب الضوء، وهو يسبب النعاس. كل أحاديات الأمين الأخرى هي منشطة أو محفزة لليقظة. الـ acetylcholine هو أيضا منشط أو محفز لليقظة.
الأصل المشترك لأحاديات الأمين
تحدثنا في مقال سابق عن “التقطع المتتالي” metamerism الذي يميز أجسام الحيوانات ثنائية الجانب bilaterian، وبينا أن هذا التقطع يظهر في الجهاز العصبي، خاصة أثناء المرحلة الجنينية الباكرة. الجهاز العصبي المركزي لدى الجنين البشري (وغير البشري) يكون مقطعا إلى قطع متتالية تسمى “القطع العصبية” neuromeres. هذه القطع العصبية تتمايز لاحقا إلى ما يلي:
- القطع النخاعية myelomeres (قطع الحبل الشوكي الذي يسمى باللاتينية “النخاع الشوكي” medulla spinalis)
- قطع الدماغ الخلفي rhombomeres
- قطع الدماغ الأوسط mesomeres
- قطع الدماغ الأمامي prosomeres
بينا في مقالات سابقة أن قطع الحبل الشوكي أو القطع النخاعية myelomeres هي الأكثر بدائية في الجهاز العصبي المركزي، واقترحنا آلية تفسر التعقيد المشاهد في القطع الدماغية. هذه الآلية هي آلية “التكتل ثم التخصص”. الخلايا تتكتل في البداية بهدف زيادة طاقتها الإنتاجية الكلية، ولكنها بعد ذلك تتخصص نوعيا.
طالما أن قطع الحبل الشوكي هي الأكثر بدائية في الجهاز العصبي المركزي فهذا يعني أن دراسة هذه القطع يمكن أن تساعدنا على فهم الشكل الأصلي للقطع الدماغية، ونحن فعلنا هذا بالفعل: في البداية تحدثنا عن مقطع الحبل الشوكي وبعد ذلك بدأنا الحديث عن المقاطع الدماغية.
الملفت في مقطع الحبل الشوكي هو أنه يخلو (حسب علمي) من خلايا مفرزة لأحاديات الأمين. مقطع الحبل الشوكي يحوي في القرنين الأمامي والجانبي خلايا تأثيرية مفرزة للـ acetylcholine، ويحوي في القرن الخلفي خلايا ربطية تفرز عددا من النواقل أهمها على ما أعتقد هو الـ glutamate (بعض الخلايا الربطية في القرن الخلفي تعتبر خلايا حسية من الرتبة الثانية second-order sensory neurons).
إذا كان الحبل الشوكي يخلو من الخلايا المفرزة لأحاديات الأمين فمن أين إذن أتت هذه الخلايا إلى الدماغ؟
أنا لا أظن أن هذه الخلايا ظهرت في الدماغ من العدم. على الأغلب أن هذه الخلايا كانت موجودة في الحبل الشوكي سابقا ولكنها اختفت منه لسبب تطوري. ما يدعم هذه الفرضية هو الصورة التالية:

الرسم الذي على اليسار يعبر عن الجهاز العصبي لدودة حلقية، والرسم الذي على اليمين يعبر عن الجهاز العصبي لفأر. الملفت في هذين الرسمين هو الفرق في توزع الخلايا المفرزة للـ serotonin؛ في الجهاز العصبي للدودة الحلقية هناك خلايا مفرزة للـ serotonin في كل قطعة عصبية، ولا يوجد تكتل لهذه الخلايا في الدماغ. في الجهاز العصبي للفأر لا يبدو أن هناك خلايا مفرزة للـ serotonin في الحبل الشوكي، وهناك تكتل لهذه الخلايا في الدماغ.
أنا أظن أن الجهاز العصبي للدودة الحلقية هو أقرب للوضع الأصلي الذي كان موجودا لدى ثنائيات الجانب البدائية urbilaterians. على ما يبدو فإن الخلايا المفرزة للـ serotonin كانت في الأصل موجودة في كل القطع العصبية المكونة للجهاز العصبي المركزي، ولكن هذه الخلايا اختفت من الحبل الشوكي عند الفقاريات (أو ربما هي ما زالت موجودة ولكن بعدد قليل جدا). في المقابل هذه الخلايا تكتلت في الدماغ وأنتجت أنوية الرفاء raphe nuclei في جذع الدماغ والخلايا المفرزة لأحاديات الأمين في ما تحت الثلم hypothalamus.
ما يبدو لي هو أن الطريقة التي تطورت بها الخلايا المفرزة لأحاديات الأمين في الدماغ تتماشى مع مبدأ “التكتل ثم التخصص”. لو نظرنا إلى جذع الدماغ فسنجد أن الخلايا المفرزة لأحاديات الأمين هي متكتلة على شكل عمود من الأنوية هو أنوية الرفاء، ولكن هذه الخلايا تفرز ناقلا واحدا هو الـ serotonin، أي أنها متكتلة ولكنها ليست متخصصة. لو صعدنا أكثر نحو الدماغ الأمامي فسنجد أن تكتل الخلايا ازداد ولكنها باتت متخصصة، بمعنى أنها لم تعد تفرز ناقلا واحدا فقط ولكنها صارت تفرز عددا من النواقل المختلفة (الـ dopamine والـ melatonin والـ histamine وربما أيضا الـ norepinephrine أو octopamine).
لو صح هذا التصور فهو قد يعني أن الخلايا المفرزة للـ serotonin هي “الأصل المشترك” للخلايا المفرزة لأحاديات الأمين المختلفة، بمعنى أن الخلايا المفرزة للـ dopamine والـ histamine إلخ هي كلها متحدرة من خلايا مفرزة للـ serotonin شبيهة بالخلايا الموجودة في أنوية الرفاء. هذا الطرح هو مجرد نظرية شخصية من عندي.
الوظيفة الأصلية للخلايا المفرزة لأحاديات الأمين
ما هي الوظيفة الأصلية للخلايا المفرزة لأحاديات الأمين؟
لو قبلنا النظرية التي طرحتها في الأعلى (والتي هي من بنات أفكاري) فهذا يعني أن الخلايا المفرزة للـ serotonin هي ربما الأصل المشترك للخلايا المفرزة لأحاديات الأمين. إذن لو عرفنا الوظيفة الأصلية للخلايا المفرزة للـ serotonin فنحن سنعرف الوظيفة الأصلية للخلايا المفرزة لأحاديات الأمين.
أنا تحدثت في مقال سابق (باختصار) عن الوظيفة الأصلية للـ serotonin لدى ثنائيات الجانب: هذا الناقل يولد في الجهاز العصبي شعور “الشبع” أو “اللذة”.
كلمة “الشبع” هي على ما أعتقد كلمة ذات معنى فلسفي معقد نسبيا ولا تصلح لوصف وظيفة فيزيولوجية في جسم حيوان بدائي. لو أردنا وصف وظيفة الـ serotonin لدى الديدان بعبارات علمية دقيقة فيجب أن نقول ما يلي: إفراز الـ serotonin في الجهاز العصبي للدودة يحصل عندما تعثر الدودة على كمية وافرة من الغذاء، وهذا الإفراز يؤدي إلى تثبيط الحركة الانتقالية inhibition of locomotion للدودة، بمعنى أن الدودة تتوقف عن الحركة الانتقالية من مكان إلى آخر.
بالإضافة إلى ذلك الـ serotonin هو محفز لليقظة (كسائر أحاديات الأمين).
المقصود بكلمة “اليقظة” arousal هو القدرة الحسية. عندما يكون الحيوان يقظا فإنه يكون أقدر على الاستقبال الحسي.
الـ acetylcholine هو أيضا محفز لليقظة، ولكنه (حسب علمي) محفز للحركة الانتقالية.
إذن الفرق الأساسي بين الـ serotonin وبين الـ acetylcholine هو أن الأول مثبط للحركة الانتقالية والثاني محفز لها.
هذا قد يكون الفرق الأصلي بين الـ serotonin والـ acetylcholine: كلا هذين الناقلين هما محفزان للاستقبال الحسي، وعلى ما أعتقد فإن كليهما محفزان للحركة الحشوية (مثلا حركة الجهاز الهضمي)، ولكن الـ serotonin هو مثبط للحركة الهيكلية أو الانتقالية، بخلاف الـ acetylcholine الذي هو محفز للحركة الهيكلية أو الانتقالية.
لو نظرنا إلى أحاديات الأمين التي يفترض أنها أحدث عمرا من الـ serotonin (كالـ dopamine والـ norepinephrine إلخ) فسنجد أنها تشترك عموما مع الـ serotonin في خاصية التحفيز الحسي أو تحفيز اليقظة، ولكن تأثيرات هذه النواقل على الحركة هي متباينة. الـ dopamine والـ norepinephrine هما محفزان للحركة الانتقالية ولكنهما مثبطان للحركة الحشوية، أي أن تأثيرهما على الحركة هو معاكس لتأثير الـ serotonin.
يرجى الانتباه إلى أن النظرية التي تقول بأن الـ serotonin هو أقدم من أحاديات الأمين الأخرى هي مجرد تخرص وأنا لا أملك دليلا عليها. من الممكن أن جميع أحاديات الأمين هي متحدرة من أصل مشترك يختلف عن الـ serotonin. لا يوجد لدي دليل على أن الـ serotonin هو أقدم من أحاديات الأمين الأخرى.
الأهم في كل ما سبق هو أن الخلايا المفرزة لأحاديات الأمين هي مرتبطة وظيفيا بالمستقبلات الحسية. أحاديات الأمين هي في الغالب محفزة للاستقبال الحسي، ولكنها أحيانا تثبطه (كما في حالة الـ melatonin الذي هو محفز للنوم). الخلايا المفرزة لأحاديات الأمين تعتمد على الإشارات الحسية الواردة من المستقبلات الحسية وترسل إشارات تأثيرية متباينة نحو الغدد والخلايا العضلية الحشوية والخلايا العضلية الهيكلية. الخلايا المفرزة لأحاديات الأمين هي إذن نوع من الخلايا العصبية الربطية التي تربط بين المستقبلات الحسية والخلايا التأثيرية (الإفرازية والتحريكية). من الممكن أن سبب تمايز الخلايا المفرزة لأحاديات الأمين الموجودة في الدماغ الأمامي هو ارتباطها بنوع (أو أكثر) من المستقبلات الحسية “الخاصة” (خاصة المستقبلات الضوئية). على سبيل المثال، من الممكن أن سبب تطور الـ melatonin هو ارتباطه بالعين الجدارية، التي هي عضو حسي خاص لا يوجد مثيل له في مناطق الجسم الأخرى. أيضا من الممكن أن الـ dopamine تطور في الأصل بسبب ارتباطه بالمستقبلات الضوئية التي أصبحت موجودة لاحقا ضمن شبكيتي العينين، ومن الممكن أن الـ histamine تطور بسبب ارتباطه بنوع خاص من المستقبلات الضوئية (أو غير الضوئية).
المقصود من هذه النظرية هو أن أحاديات الأمين تكاثرت وتمايزت بسبب ارتباطها بمستقبلات حسية خاصة من أنواع مختلفة، خاصة المستقبلات الضوئية. هذه النظرية تفسر وجود الخلايا المفرزة لأحاديات الأمين بكميات كبيرة وبأنواع متعددة في الدماغ الأمامي، وخاصة في الدماغ البيني الذي هو مرتبط تطوريا بحس البصر.
الدماغ البشري (9)

الدماغ الأوسط (midbrain) mesencephalon هو أصغر أجزاء الدماغ البشري. طول هذا الدماغ لا يتجاوز 2 سم، وقطره يبلغ وسطيا 1.8 سم.
الدماغ البشري (8)
هذا المقال سيكون جافا نوعا ما لأنني سأتحدث فيه عن التكوين التشريحي للجسر pons ولن أتطرق كثيرا إلى الوظائف والآليات الفيزيولجية. سوف أؤجل الحديث عن الآليات والوظائف والمنعكسات إلى أن أنهي الحديث عن التكوين التشريحي لجذع الدماغ.
في الجزء السادس من هذه السلسلة تحدثت عن مقاطع النخاع المستطيل المنقاري rostral medulla oblongata. أهم ما يميز هذه المقاطع هو انتفاخ القناة المركزية central canal وتحولها إلى البطين الرابع fourth ventricle الذي يفصل بين جذع الدماغ والمخيخ cerebellum.


الآن سنتحدث عن مقطع الجسر pons. أهم ما يميز الجسر هو وجود المخيخ على ظهره. المخيخ من حيث الأصل الجنيني هو مشتق من الصفيحتين الجناحيتينalar plates للأنبوب العصبي neural tube.
ذكرنا في السابق أن الجهاز العصبي المركزي في بداية تكوينه (لدى الجنين) يكون له شكل أنبوب يسمى “الأنبوب العصبي” neural tube. لو أخذنا مقطعا عرضيا في الأنبوب العصبي فسنجد أنه يحوي في مركزه تجويفا أو قناة (ولهذا السبب سمي أنبوبا). هذه القناة تتحول إلى “القناة المركزية” central canal التي نراها لدى الإنسان البالغ في وسط الحبل الشوكي. حول القناة المركزية توجد كتل من المادة الرمادية (أجسام الخلايا العصبية). عند الجنين هذه المادة الرمادية تقسم إلى صفائح قاعدية basal plates وصفائح جناحية alar plates.

الصفيحتان القاعديتان basal plates توجدان في الناحية البطنية ventral من الأنبوب العصبي، والصفيحتان الجناحيتان alar plates توجدان في الناحية الظهرية dorsal من الأنبوب العصبي. الصفيحتان القاعديتان تتحولان في الحبل الشوكي إلى القرنين البطنيين ventral horns (الذين يحويان خلايا تأثيرية effector)، والصفيحتان الجناحيتان تتحولان إلى القرنين الظهريين dorsal horns (الذين يحويان خلايا حسية أو استقبالية).
القناة المركزية تقترب بشكل تدريجي من الجدار الظهري للنخاع المستطيل كلما اتجهنا من الجهة الذيلية caudal نحو الجهة المنقارية rostral (أي كلما اتجهنا من الأسفل إلى الأعلى)، ونفس الأمر ينطبق أيضا على كتل المادة الرمادية (سواء كانت تأثيرية أم حسية). عندما تحدثنا عن مقاطع النخاع المستطيل بينا أن المادة الرمادية في هذه المقاطع تتكتل في الناحية الظهرية، وأما الناحية البطنية فهي مكونة بمعظمها من مادة بيضاء (طرق عصبية أو ألياف عصبية).
القناة المركزية تتسع بشكل تدريجي في النخاع المستطيل كلما اتجهنا نحو الجهة المنقارية، إلى أن تتحول في النهاية إلى البطين الرابع. الاتساع في القناة المركزية يترافق مع تغير في التوزع “الطوبوغرافي” لأنوية المادة الرمادية، بمعنى أن الأنوية التأثيرية تصبح على الجهة الإنسية medial بعدما كانت تقع على الجهة البطنية ventral في مقطع الحبل الشوكي، والأنوية الحسية تصبح على الجهة الطرفية أو الوحشية lateral بعدما كانت تقع على الجهة الظهرية dorsal في مقطع الحبل الشوكي.
في المقالات الأخيرة بينا الفرق بين الحس العام general والحس الخاص special، وذكرنا أن الحس الخاص هو موجود حصرا في الدماغ. من أمثلة الحس الخاص حسا الاتزان والسمع اللذين تحدثنا عنهما في المقال الأخير. أنوية هذين الحسين تقع في منطقة الاتصال بين النخاع المستطيل المنقاري والجسر. هذه الأنوية توجد في أقصى الجهة الطرفية أو الوحشية lateral للمادة الرمادية (أنوية حس السمع توجد على الجهة الوحشية لأنوية حس الاتزان). هذا يعني أن هذه الأنوية هي مشتقة جنينيا من القسم الظهري من الصفيحتين الجناحيتين alar plates.
الجسر والمخيخ هما مشتقان جنينيا من الدماغ التالي metencephalon.

الدماغ الخلفي rhombencephalon ينقسم إلى:
- الدماغ النخاعيmyelencephalon (يتحول إلى النخاع المستطيل)
- الدماغ التالي metencephalon (يتحول إلى الجسر والمخيخ)
المخيخ هو مشتق من الصفيحتين الجناحيتين للدماغ التالي، وهو يقع على الجهة الظهرية للبطين الرابع.
بما أن المخيخ مشتق من الناحية الظهرية للصفيحتين الجناحيتين فهذا يوحي بأن المخيخ هو عضو حسي خاص، ولكن الكتب والمراجع التقليدية تصنف المخيخ على أنه عضو تحريكي motor.
الجسر يقع على الجهة البطنية للبطين الرابع. تكوين الجسر يشبه تكوين النخاع المستطيل (الجسر هو في الحقيقة مجرد امتداد وظيفي للنخاع المستطيل). ما يلي مقطع في الجسر الذيلي:

هذا المقطع يشمل كلا من الجسر والمخيخ. المناطق التي تقع فوق الخطين الأحمرين هي في المخيخ وليست في الجسر. مقطع الجسر الذيلي الذي سنتحدث عنه هو فقط المناطق الواقعة تحت الخطين الأحمرين.
تكوين مقطع الجسر يشبه تكوين مقطع النخاع المستطيل. المنطقة البطنية (السفلية) من المقطع تحوي كمية أكبر من المادة البيضاء، والمنطقة الظهرية (العلوية) من المقطع (التي تقع مباشرة تحت قعر البطين الرابع) تحوي كمية أكبر من المادة الرمادية.
الجسر القاعدي
على الوجه البطني للجسر توجد كتلتان بارزتان تشبهان الهرمين pyramids الموجودين على الوجه البطني للنخاع المستطيل ولكنهما ذواتا امتداد أعرض. هاتان الكتلتان تسميان “الجسر القاعدي” basilar pons.

الجسر القاعدي يحوي طرقا نازلة من قشرة المخ:
- الطريق القشري الشوكي corticospinal tract (نحو قرون الحبل الشوكي)
- الطريق القشري البصلي corticobulbar tract (نحو أنوية الأعصاب القحفية)
هذه الطرق تسمى “الطرق الهرمية” pyramidal tracts أو “الجهاز الهرمي” pyramidal system، لأنها عندما تصل إلى النخاع المستطيل تكون الهرمين pyramids. هذه الطرق هي مسؤولة عن التحكم الواعي أو الإرادي بالأعصاب الشوكية والقحفية.
إلى جانب الطرق الهرمية هناك في الجسر القاعدي ألياف نازلة أخرى تسمى الألياف القشرية الجسرية corticopontine fibers. هذه الألياف لا تصل إلى الهرمين ولكنها تتشابك مع أنوية ربطية موجودة في الجسر القاعدي تسمى الأنوية الجسرية pontine nuclei.

أصل الأنوية الجسرية pontine nuclei يشبه أصل الأنوية الزيتونية السفلية inferior olivary nuclei الموجودة داخل الزيتونتين olives الظاهرتين على جانبي الهرمين في النخاع المستطيل—الأنوية الجسرية هي مشتقة من الصفيحتين الجناحيتين alar plates للدماغ التالي metencephalon، والأنوية الزيتونية السفلية هي مشتقة من الصفيحتين الجناحيتين للدماغ النخاعي myelencephalon.

الأنوية الزيتونية السفلية والأنوية الجسرية هي مشتقة من الصفيحتين الجناحيتين رغم كونها تقع في الجهة البطنية من جذع الدماغ. الملفت هو أن الألياف الصادرة من الأنوية الزيتونية السفلية والأنوية الجسرية تذهب نحو المخيخ. الألياف الصادرة من الأنوية الزيتونية السفلية تسمى الألياف الزيتونية المخيخية olivocerebellar fibers وهي تذهب إلى المخيخ عبر السُّوَيقة المخيخية السفلية inferior cerebellar peduncle (التي تصدر من النخاع المستطيل المنقاري). الألياف الصادرة من الأنوية الجسرية تسمى الألياف الجسرية المخيخية pontocerebellar fibers وهي تذهب إلى المخيخ عبر السُّوَيقة المخيخية الوسطى middle cerebellar peduncle (التي تصدر من الجسر). الألياف الجسرية المخيخية تظهر بشكل معترض في مقطع الجسر. وجود هذه الألياف هو سبب الشكل العريض للجسر القاعدي مقارنة مع هرمي النخاع المستطيل.



خلاصة ما سبق هي أن الجسر القاعدي basilar pons يتكون من ألياف طولانية نازلة هي الألياف القشرية الشوكيةcorticospinal والقشرية البصلية corticobulbar والقشرية الجسرية corticopontine. الألياف القشرية الشوكية وبعض الألياف القشرية البصلية تنزل نحو النخاع المستطيل وتكون الهرمين. الألياف القشرية الجسرية تتشابك مع الأنوية الجسرية الموجود في الجسر القاعدي (أظن أنها تسمى أيضا “الأنوية الجسرية العميقة” deep pontine nuclei). الأنوية الجسرية (العميقة) تصدر أليافا معترضة تسمى الألياف الجسرية المخيخية pontocerebellar (تسمى أيضا “الألياف الجسرية المعترضة” transverse pontine fibers). هذه الألياف المعترضة تذهب نحو المخيخ عبر السُّوَيقة المخيخية الوسطى.
الغطاء
كلمة tegmentum هي تسمية تطلق على مناطق الجسر التي تقع خلف الجسر القاعدي (على جهته الظهرية)، أي أنها تشمل كل ما يقع بين الجسر القاعدي وقعر البطين الرابع.
في القاموس الطبي السوري كلمة tegmentum ترجمت إلى “السَّقِيْفَة”. أنا رجعت إلى أربعة أو خمسة قواميس لاتينية ولم أتمكن من إيجاد الأصل الذي ترجمه مخترعو اللغة الطبية السورية إلى “السقيفة”. على ما يبدو فإن هذه الترجمة لا أصل لها وهي مجرد اختراع من العدم (وهذا ليس شيئا نادرا في اللغة الطبية السورية).
كلمة tegumentum أوtegmentum أو tegimentum هي مأخوذة من الفعل tegere الذي يعني “التغطية” to cover. الجذر *teg- أو *steg- هو هندو-أوروبي الأصل وهو مصدر الكلمة اليونانية القديمة τέγος التي تعني “سقف”. هذا الرابط فيه إحصاء للمواضع التي وردت فيها كلمة tegumentum في الكتب اللاتينية القديمة. في معظم الأحيان هذه الكلمة وردت بمعنى “غطاء” a covering, cover، ولكنها وردت أيضا بمعنى “درع” armor أو “حماية” protection.
في علم البيولوجيا (المعاصر) كلمة tegument تستخدم كمرادف لكلمة integument. معنى هاتين الكلمتين هو “طبقة خارجية تغطي جزءا من الحيوان أو النبات”. ما يلي هو تعريف الكلمة في قاموس Oxford:
tegument
Pronunciation: /ˈtɛgjʊm(ə)nt/
noun
chiefly Zoology
the integument of an organism, especially a parasitic flatworm.
Origin:
late Middle English (in the general sense ‘a covering or coating’): from Latin tegumentum, from tegere ‘to cover’
ما يلي هو تعريف كلمة integument:
integument
Pronunciation: /ɪnˈtɛgjʊm(ə)nt/
noun
a tough outer protective layer, especially that of an animal or plant:
this chemical compound is found in the integument of the seed
Origin:
early 17th century (denoting a covering or coating): from Latin integumentum, from the verb integere, from in- ‘in’ + tegere ‘to cover’
نفس التعاريف موجودة في قاموس Encarta:
tegument
teg·u·ment [téggyəmənt]
(plural teg·u·ments)
noun
integument: the protective outer covering of an organism
[15th century. < Latin tegumentum “covering” < tegere “to cover”]
________________________________________________________________________
integument
in·teg·u·ment [in téggyəmənt]
(plural in·teg·u·ments)
noun
protective layer: an outer protective layer or part of an animal or plant, e.g. a shell, rind, husk, or skin
[Early 17th century. < Latin integumentum < integere “cover up” < tegere “to cover”]
الخلاصة هي أن كلمةtegumentum في اللغة اللاتينية الكلاسيكية (القديمة) تعني “غطاء” أو “درع” أو “حماية”، وفي اللغة اللاتينية العلمية (المعاصرة) تعني “طبقة خارجية تغطي جزءا من الحيوان أو النبات” (مثلا كالقشرة التي تغطي الديدان أو بذور النبات، أو كالصدفة التي تغطي بعض الحيوانات إلخ).
السؤال الكبير الذي يطرح نفسه هو من أين أتى القاموس الطبي السوري بكلمة “السقيفة” كترجمة لـ tegmentum؟ هذه الترجمة ليس لها أساس في اللغة اللاتينية القديمة ولا في اللغة اللاتينية المعاصرة. هي مجرد ترجمة خاطئة. هذا مجرد مثال على الترجمات الخاطئة التي ترد في قاموس اللغة الطبية السورية.
في هذا المقال سوف أترجم كلمة tegmentum إلى “غطاء”. هذه الترجمة هي بسيطة وفي نفس الوقت صحيحة. طبعا القارئ (الجاد) يجب أن يضع في رأسه كلمة tegmentum لأن هذه الكلمة هي المصطلح العلمي الحقيقي، وأما الترجمات العربية فهي ليست مصطلحات علمية وليست قريبة من أن تكون مصطلحات علمية (إلا لدى الأشخاص الذين لا يفهمون أصلا ما هو معنى كلمة مصطلح علمي).
الغطاء tegmentum هو كما قلنا جميع مناطق الجسر التي تقع بين الجسر القاعدي وقعر البطين الرابع. القسم البطني من الغطاء يحوي عددا من الطرق العصبية الصاعدة. من هذه الطرق ما يلي:
- الشريط الإنسي medial lemniscus الذي ينقل إشارات حس الاهتزاز والحس الذاتي من النواة النحيلة nucleus gracilis والنواة الوتدية nucleus cuneatus (في النخاع المستطيل الذيلي)، واللتان بدورهما استقبلتا هذه الأحاسيس من العمودين الظهرين dorsal columns (القادمين من الحبل الشوكي).
- الطريق الشوكي الثلميspinothalamic tract (أو الشريط الشوكي spinal lemniscus) الذي ينقل إشارات حس الألم والحرارة من الحبل الشوكي.
- الطريق السمعي الذي يبدأ من النواتين القوقعيتين cochlear nuclei (تمتدان بين النخاع المستطيل المنقاري والجسر الذيلي) وتتشابك بعض أليافه في الجسر الذيلي مع الجسم شبه المنحرف trapezoid body والأنوية الزيتونية العلوية superior olivary nuclei، ثم تصعد هذه الألياف عبر الشريط الوحشي lateral lemniscus نحو التليلة السفليةinferior colliculus في سقف الدماغ الأوسط midbrain tectum.

الأنوية الزيتونية العلوية superior olivary nuclei هي غير ظاهرة في مقطعي الجسر الذيلي الذين أوردناهما في الأعلى. هذه الأنوية هي امتداد منقاري للأنوية الزيتونية السفلية، ولكنها على ما أظن ليست موجودة ضمن الزيتونتين olives لأن الزيتونتين تنتهيان عند الحدود المنقارية للنخاع المستطيل. حسب ما يرد في المصادر فإن الأصل الجنيني للأنوية الزيتونية العلوية هو من الصفيحة القاعدية basal plate (ولكن أحد المصادر القديمة نسبيا طرح احتمال أن يكون أصلها من الصفيحة الجناحية). الأنوية الزيتونية العلوية ترسل أليافا صاعدة نحو التليلة السفليةinferior colliculus في سقف الدماغ الأوسط. إذن الأنوية الزيتونية العلوية -بخلاف الأنوية الزيتونية السفلية- هي ليست مرتبطة بشكل مباشر بالمخيخ، لا من حيث الأصل الجنيني ولا من حيث التشابك العصبي.
الغطاء tegmentum يحوي أيضا طرقا عصبية تضم أليافا صاعدة ونازلة. من هذه الطرق:
- الطريق الغطائي المركزي central tegmental tract
- الحزمة الطولانية الإنسية medial longitudinal fasciculus (اختصارا MLF)
هذان الطريقان يمتدان على طول جذع الدماغ (من الدماغ الأوسط midbrain وحتى النخاع المستطيل). أنا تحدثت في مقالات سابقة عن بعض الألياف التي تمر عبر هذين الطريقين. مثلا “الطريق الغطائي المركزي” يحوي ألياف “الطريق المفرد الثلمي” solitariothalamic tract الذي ينقل حس الذوق نحو الثلم thalamus من القسم المنقاري لنواة الطريق المفرد nucleus of the solitary tract. سوف أؤجل الحديث عن بقية الألياف المارة في هذين الطريقين إلى مقال لاحق.
بالنسبة للمادة الرمادية في الغطاء tegmentum فهي تنقسم عموما إلى ثلاثة أقسام (كما في النخاع المستطيل):
- أنوية قحفية (أنوية الأعصاب القحفية رقم 8 و7 و6 و5)
- أنوية شبكية (في التشكل الشبكي reticular formation)
- أنوية ربطية (مثلا الجسم شبه المنحرف والأنوية الزيتونية العلوية)
الأنوية القحفية

الأنوية القحفية تتوزع من الجهة الجانبية أو الوحشية lateral إلى الجهة الإنسية medial على النحو التالي:
- في أقصى الجهة الوحشية توجد أنوية الحس الجسماني الخاصspecial somatic ، أي الأنوية القوقعية والدهليزية التي تستقبل حسي السمع والاتزان عبر العصب القحفي رقم 8. هذه الأنوية توجد في الجسر الذيلي (ولها امتداد أيضا في النخاع المستطيل المنقاري).
- بعد ذلك توجد نواة الحس الجسماني العام general somatic، التي هي النواة الحسية للعصب القحفي رقم 5 trigeminal sensory nucleus. هذه النواة تستقبل الحس الجسماني العام القادم من الوجه عبر العصب القحفي رقم 5 (العصب ثلاثي التوائم trigeminal nerve). هذه النواة تمتد على طول جذع الدماغ من النخاع المستطيل وحتى الدماغ الأوسط.
- ثم تأتي نواة الحس الحشويvisceral ، التي هي نواة الطريق المفرد nucleus of the solitary tract. القسم المنقاري من هذه النواة (موجود في الجسر الذيلي والنخاع المستطيل المنقاري) يستقبل حس الذوق (حس حشوي خاصspecial visceral ) عبر العصب القحفي رقم 7 (في الجسر الذيلي) والعصب القحفي رقم 9 (في النخاع المستطيل المنقاري).
- على الجهة الإنسية medial للنواة السابقة توجد النواة التأثيرية الحشوية visceral effector (جانب المتعاطفة parasympathetic)، التي هي في الجسر الذيلي النواة اللعابية العلوية superior salivatory nucleus. هذه النواة هي مسؤولة عن إفراز اللعاب والدمع عبر العصب القحفي رقم 7 (العصب الوجهي facial nerve). هذه النواة يمكن اعتبارها امتدادا منقاريا للنواة اللعابية السفلية inferior salivatory nucleus الموجودة في النخاع المستطيل المنقاري والمسؤولة عن إفراز اللعاب عبر العصب القحفي رقم 9. النواة اللعابية السفلية بدورها يمكن اعتبارها امتدادا منقاريا للنواة الظهرية للعصب المبهم dorsal nucleus of the vagus nerve المسؤولة عن التعصيب جانب المتعاطف parasympathetic لكل أحشاء الجسم ما عدا القلب والأحشاء الحوضية.
- في الجهة الإنسية توجد نواة محركة جسمانية somatic motor هي نواة العصب القحفي رقم 6 المسؤولة عن تحريك العين نحو الاتجاه الوحشي lateral. هذه النواة توجد في الجسر الذيلي.
- النواة المحركة للعصب القحفي رقم 7 (التي تسمى “النواة المحركة الوجهية” facial motor nucleus) هي مسؤولة عن تحريك عضلات الوجه (التعابير الوجهية). هذه النواة هي “حشوية جسمانية” somatovisceral أو “تأثيرية حشوية خاصة” special visceral efferent ويمكن اعتبارها امتدادا نحو الجسر الذيلي للنواة المبهمة nucleus ambiguus الموجودة في النخاع المستطيل والمسؤولة عن تحريك عضلات الحنجرة والبلعوم عبر العصبين القحفيين رقم 9 و10. نواة العصب القحفي رقم 7 لها امتداد منقاري بعيد هو النواة المحركة للعصب القحفي رقم 5 (التي تسمى “النواة المحركة ثلاثية التوائم” trigeminal motor nucleus) الموجودة في وسط الجسر. هذه النواة هي مسؤولة عن تحريك عضلات المضغ muscles of mastication.
من يقارن توزع الأنوية القحفية في الجسر مع توزع الأنوية القحفية في النخاع المستطيل سوف يرى أن هذه الأنوية تشكل ما يشبه الأعمدة الوظيفية المتقطعة. مثلا النواة المبهمة nucleus ambiguus (في النخاع المستطيل) والنواة المحركة الوجهية facial motor nucleus (في أسفل الجسر) والنواة المحركة ثلاثية التوائم trigeminal motor nucleus (في وسط الجسر) تشكل عمودا محركا “حشويا جسمانيا” somatovisceral motor. هذا العمود هو متقطع إلى أنوية ولكن الأنوية تؤدي وظائف متشابهة. نفس الأمر ينطبق على الأنوية التأثيرية الحشوية visceral effector (جانب المتعاطفة parasympathetic)، ونفس الأمر ينطبق ربما على الأنوية المحركة الجسمانية somatic motor التي تشمل نواة العصب رقم 12 (في النخاع المستطيل) ونواة العصب القحفي رقم 6 (في الجسر) ونواتي العصبين القحفيين رقم 4 و3 (في الدماغ الأوسط).
بالنسبة للأعمدة الحسية فهي أقل تقطعا وأكثر وضوحا. نواة الطريق المفرد nucleus of the solitary tract تعبر عن عمود حسي حشوي visceral sensory يمتد من النخاع المستطيل إلى الجسر الذيلي، والنواة الحسية للعصب القحفي رقم 5 هي عمود حسي جسماني somatic sensory يمتد على طول جذع الدماغ ويتمادى في الأسفل مع القرن الظهري للحبل الشوكي.
النواة الحسية للعصب القحفي رقم 5 هي متقطعة إلى ثلاث أنوية:
- النواة الشوكية spinal nucleus (في النخاع المستطيل) تستقبل من الوجه حس الألم والحرارة واللمس غير الدقيق crude touch.
- النواة الحسية الرئيسية principal sensory nucleus (في الجسر) تستقبل من الوجه حس اللمس الدقيق fine touch والاهتزاز والحس الذاتي الواعي conscious proprioception.
- نواة الدماغ الأوسط mesencephalic nucleus (في الدماغ الأوسط) تستقبل من الوجه ربما الحس الذاتي غير الواعي unconscious proprioception.

الجدول التالي يبين جميع الأنوية القحفية الموجودة في جذع الدماغ مصنفة حسب وظائفها وحسب الأعصاب القحفية المتصلة معها. العصبان القحفيان الأول والثاني هما مستثنيان لأنهما لا يصدران من جذع الدماغ.
| Effector | Receptor | ||||||
| somatic | somatovisceral | visceral | visceral | general somatic | special somatic | ||
| CN III | oculomotor nucleus | Edinger–Westphal nucleus | |||||
| CN IV | trochlear nucleus | ||||||
| CN V | trigeminal motor nucleus | trigeminal sensory nucleus | |||||
| CN VI | abducens nucleus | ||||||
| CN VII | facial motor nucleus | superior salivatory nucleus | solitary nucleus | trigeminal sensory nucleus | |||
| CN VIII | vestibular nuclei | cochlear nuclei | |||||
| CN IX | nucleus ambiguus | inferior salivatory nucleus | solitary nucleus | trigeminal sensory nucleus | |||
| CN X | nucleus ambiguus | dorsal nucleus of the vagus | solitary nucleus | trigeminal sensory nucleus | |||
| CN XII | hypoglossal nucleus | ||||||
الأنوية الشبكية
توزع الأنوية الشبكية في الجسر يشبه التوزع الذي بيناه سابقا في النخاع المستطيل. الأنوية التأثيرية (كبيرة الخلاياmagnocellular ) توجد في الجهة الإنسية، والأنوية الاستقبالية (صغيرة الخلايا parvocellular) توجد في الجهة الوحشية.
الأنوية الشبكية الاستقبالية أو الإيرادية afferent في النخاع المستطيل والجسر تشمل ما يلي (من الأسفل إلى الأعلى):
- النواة الشبكية الجانبية lateral reticular nucleus (في النخاع المستطيل)
- المنطقة الشبكية صغيرة الخلايا parvocellular reticular area (في النخاع المستطيل المنقاري والجسر الذيلي)
- المنطقة جانب الذراعية parabrachial area (في الجسر المنقاري)
- النواة السويقية الجسرية pedunculopontine nucleus والنواة الغطائية الجانبية الظهرية laterodorsal tegmental nucleus
النواتان الأخيرتان تعتبران أحيانا نواة واحدة اسمها “النواة السويقية الجسرية” pedunculopontine nucleus. اسم هذه النواة يدل على أنها تقع في الجسر، ولكن الرسومات تظهرها في الدماغ الأوسط.
الأنوية الشبكية التأثيرية أو الإصدارية efferent في النخاع المستطيل والجسر تشمل ما يلي (من الأسفل إلى الأعلى):
- النواة الشبكية عملاقة الخلايا gigantocellular reticular nucleus (في النخاع المستطيل)
- النواة الشبكية جانب المحورية paramedian reticular nucleus (في النخاع المستطيل)
- النواة الشبكية الجسرية الذيلية caudal pontine reticular nucleus (في الجسر الذيلي)
- النواة الشبكية الجسرية الفموية oral pontine reticular nucleus (في الجسر المنقاري)
- النواة الشبكية الغطائية الجسرية pontine reticulotegmental nucleus (في الجسر المنقاري)
- الموضع الأزرق locus caeruleus (في الجسر المنقاري)
بعض الأنوية الشبكية سالفة الذكر تسمى “قبل مخيخية” precerebellar لأنها تصدر أليافا عصبية نحو المخيخ. هناك ثلاثة أنوية شبكية “قبل مخيخية” هي:
- النواة الشبكية الجانبية lateral reticular nucleus (في النخاع المستطيل)
- النواة الشبكية جانب المحورية paramedian reticular nucleus (في النخاع المستطيل)
- النواة الشبكية الغطائية الجسرية pontine reticulotegmental nucleus (في الجسر المنقاري)
النواة الشبكية الجانبية lateral reticular nucleus تشبه النواة الزيتونية السفلية inferior olivary nucleus من حيث كونها نواة استقبالية تستقبل أليافا من مصدرين:
- ألياف صاعدة من الحبل الشوكي
- وألياف نازلة من النواة الحمراء red nucleus في الدماغ الأوسط
النواة الشبكية الجانبية تشبه أيضا النواة الزيتونية السفلية من حيث موقعها في مقطع النخاع المستطيل، ومن حيث كونها تصدر أليافا نحو المخيخ. الفرق الأساسي بين النواتين (حسب ما يبدو لي) هو في نوعية الألياف الشوكية الصاعدة التي تستقبلها كل منهما. النواة الزيتونية السفلية تستقبل ألياف الطريق الشوكي الزيتوني spinoolivary tract التي تنقل الحس الذاتي، والنواة الشبكية الجانبية تستقبل ألياف الطريق الشوكي الشبكي spinoreticular tract التي تنقل حس الألم والحرارة. بناء على ما سبق يمكننا أن نتخيل أن النواة الشبكية الجانبية تؤدي وظيفة مناظرة لوظيفة النواة الزيتونية السفلية (أنا لم أتحدث بعد عن وظيفة النواة الزيتونية السفلية ولكن ما يبدو من بعض الدراسات الحديثة هو أن وظيفة هذه النواة هي تصحيح أو تحديث المعلومات المخزنة في المخيخ بناء على المعطيات الحسية الواصلة إليها).
حسب ما يرد في بعض المصادر فإن النواة الشبكية الجانبية ترسل أيضا أليافا نحو الأنوية الشبكية التأثيرية الموجودة في القسم الإنسي medial من التشكل الشبكي. أنا في مقال سابق قارنت بين هذه الوظيفة وبين وظيفة الأنوية الدهليزية التي تستقبل البيانات الحسية الميكانيكية من الحبل الشوكي والأذن الداخلية وتستخدم هذه البيانات للتدخل في منعكسات التوتر العضلي muscle tone التي تديرها الأنوية الشبكية التأثيرية.
بما أن النواة الشبكية الجانبية ترسل أليافها نحو جهتين (نحو المخيخ ونحو الأنوية الشبكية التأثيرية) فهذا يوحي (مع خط تحت كلمة يوحي) بأنها تؤدي وظيفتين: الوظيفة الأولى تشبه وظيفة النواة الزيتونية السفلية (تحديث معلومات المخيخ)، والوظيفة الثانية تشبه وظيفة الأنوية الدهليزية (تعديل التوتر العضلي).
بالنسبة للنواة الشبكية جانب المحورية paramedian reticular nucleus والنواة الشبكية الغطائية الجسرية pontine reticulotegmental nucleus فهاتان النواتان لهما قصة مختلفة عن قصة النواة الشبكية الجانبية. هاتان النواتان هما على ما يبدو نواتان تأثيريتان وليستا استقباليتين، بدليل موقعهما في مقاطع جذع الدماغ: هما تقعان على الجهة الإنسية للأنوية الشبكية التأثيرية. هاتان النواتان لا تستقبلان إشارات حسية على ما يبدو ولكنهما تستقبلان إشارات قادمة (عبر الحزمة الطولانية الإنسية MLF) من سقف الدماغ الأوسط ومن الأنوية الدهليزية وتستخدمانها لتنسيق المنعكسات المركبة المتعلقة بعملية الرؤية (وربما السمع أيضا لدى بعض الحيوانات) والتي سأتحدث عن بعضها لاحقا. وظيفة هاتين النواتين إذن هي التنسيق الحركي motor coordination. هذه الوظيفة تشبه وظيفة “القشرة قبل المحركة” premotor cortex في المخ. الألياف التي تصدر من هاتين النواتين وتتجه نحو المخيخ هي ربما شبيهة بالألياف التي تنزل من القشرة قبل المحركة نحو المخيخ عبر الأنوية الجسرية pontine nuclei.
وظيفة الألياف التي تنزل نحو المخيخ من القشرة “قبل المحركة” premotor (والقشرة المحركة motor) هي ليست واضحة، ولكنني أظن (بناء على فهمي لبعض النظريات والطروحات) أن وظيفة هذه الألياف هي تحفيز المخيخ لإرسال نسخة معدلة من الإشارات التحريكية التي أرسلت إليه. المخيخ حسب بعض الطروحات الحديثة هو في الأساس عضو ذاكرة، بمعنى أنه يقوم بتخزين الإشارات العصبية الواصلة إليه في القشرة المخيخية cerebellar cortex. القشرة المخيخية تحوي على ما يبدو “خريطة تحريكية” لعضلات الجسم ضمن ما يسمى بطبقة خلايا بوركينيي Purkinje cell layer. هذه “الخريطة التحريكية” هي مرتبطة مع “خريطة حسية” موجودة ضمن الطبقات الأخرى للقشرة المخيخية. الهدف من الربط بين الخريطة التحريكية والخريطة الحسية هو زيادة دقة الخريطة التحريكية. الخلايا الدماغية التي تولد إشارات تحريكية ترسل نسخة من هذه الإشارات إلى المخيخ بهدف تعديلها بناء على الخريطة التحريكية الموجودة فيه.
الوظائف التحريكية للأنوية الشبكية
الأنوية الشبكية هي مسؤولة عن وظائف أو منعكسات عديدة. في مقال سابق تحدثنا عن إحدى الوظائف التحريكية البدائية لهذه الأنوية وهي وظيفة تقليص وإرخاء العضلات الباسطة المحورية axial extensor muscles (العضلات المضادة للجاذبية anti-gravity muscles). هذه الوظيفة كانت في الأصل تهدف (على ما أظن) لتحريك الجسم، ولكنها لاحقا أصبحت تهدف للحفاظ على انتصاب وتوازن الجسم (ما يسمى posture) بالاعتماد على ما يسمى بالتوتر العضلي muscle tone. حسب كثير من المصادر فإن الأنوية الشبكية المسؤولة عن هذه الوظيفة هي أساسا الأنوية الموجودة في النخاع المستطيل (تثبيطية) وفي الجسر (إثارية).
الوظيفة التحريكية الثانية للأنوية الشبكية التي تحدثنا عنها هي وظيفة التحكم بحركة العينين بهدف تحسين جودة الاستقبال الحسي عبرهما. سوف أذكر المزيد عن هذه الوظيفة في مقال لاحق (بعد أن أتحدث عن تكوين الدماغ الأوسط).
هناك وظيفة تحريكية ثالثة تحدثنا عنها (في هذا المقال) هي وظيفة تنظيم التنفس وإيقاعه. الأنوية الشبكية المحفزة للشهيق والزفير توجد في النخاع المستطيل، والأنوية المنظمة لإيقاع التنفس respiratory rhythm توجد ربما في الجسر.

أحد المراكز الذي يعتقد أنه مسؤول عن تنظيم إيقاع التنفس هو المركز المسمى “المركز المنظم للتنفس” pneumotaxic center (كلمة (táxis) τάξις في اليونانية تعني “تنظيم”) أو “المجموعة التنفسية الجسرية” pontine respiratory group. هذا المركز هو موجود ضمن الأنوية الشبكية التي تسمى “المجمّع جانب الذراعي” parabrachial complex. هذا المجمع هو موجود في الجسر المنقاري بجانب السويقة المخيخية العلوية (السويقة المخيخية العلوية تسمى أيضا “الذراع الضامة” brachium conjunctivum، ومن هنا جاءت تسمية المجمّع جانب الذراعي). أنوية المجمع جانب الذراعي هي متصلة مع نواة الطريق المفرد nucleus of the solitary tract عبر العديد من الألياف العصبية، وهذا يفسر دور المجمع في تنظيم إيقاع التنفس. المجمع يستقبل أيضا ألياف حس الذوق من القسم المنقاري لنواة الطريق المفرد ويوصل هذا الحس إلى الثلم thalamus عبر ألياف صاعدة.
الخلاصة هي أن “المجمّع جانب الذراعي” parabrachial complex هو على ما يبدو مسؤول عن تنظيم عدد من المنعكسات أو الوظائف المتعلقة بالحس الحشوي visceral الوارد إلى نواة الطريق المفرد.
النواة الاستقبالية المسماة “النواة الشبكية صغيرة الخلايا” parvocellular reticular nucleus تلعب أيضا دورا في التنفس. تحفيز القسم الذيلي من هذه النواة يحفز الزفير، وهناك اتصالات بين قسمها المنقاري وبين المجمع جانب الذراعي. هناك أيضا اتصالات بين هذه النواة وبين النواة ثلاثية التوائم في الدماغ الأوسط mesencephalic trigeminal nucleus التي تستقبل الحس الذاتي (غير الواعي؟) من العصب القحفي الخامس. النواة الشبكية صغيرة الخلايا هي متصلة أيضا مع نواة العصب القحفي السابع (العصب الوجهي facial) المسؤول عن تحريك عضلات الوجه، ومع نواة العصب القحفي الثاني عشر (العصب تحت اللساني hypoglossal) المسؤول عن تحريك اللسان. هذه الاتصالات توحي بأن النواة الشبكية صغيرة الخلايا تستقبل حسا جسمانيا وتساهم في تنظيم منعكسات جسمانية أو “جسمانية حشوية”.
بالنسبة للموضع الأزرق locus caeruleus والنواة السويقية الجسرية pedunculopontine nucleus والأنوية الشبكية الموجودة في الدماغ الأوسط فهذه كلها تلعب دورا في تنظيم اليقظة arousal أو التنشيط activation. هناك اعتقاد لدى بعض الباحثين بأن هذه الأنوية هي مسؤولة أيضا عن توليد بعض المنعكسات الحركية الإيقاعية (كالمشي والركض)، ولكن هذا الاعتقاد هو محل جدل.
في المقال القادم سوف أتحدث عن تكوين الدماغ الأوسط، وسوف أتحدث عن بعض المنعكسات والآليات التي ينظمها هذا الدماغ.

{kind=link}
{kind=link}