كثير من الكائنات الحية تملك القدرة على الحركة. بعض الكائنات الحية تستطيع أن تحرك أجزاء من جسمها، وبعضها تستطيع أن تنقل جسمها بكامله من مكان إلى آخر. الحركة التي تؤدي لنقل الجسم بكامله من مكان إلى آخر تسمى الحركة الانتقالية locomotion.الحركة الانتقالية هي خاصية بيولوجية بدائية جدا، بدليل انتشارها الواسع لدى الكائنات الحية أحادية الخلية حقيقية النواة eukaryotes.الباحث السويدي Carolus Linnaeus (في القرن 18) قسم الكائنات الحية إلى مملكتين:
- مملكة الحيوان Regnum Animale
- مملكة النبات Regnum Vegetabile
هذا التقسيم لم يفرد مملكة خاصة للكائنات الحية المجهرية أحادية الخلية التي اكتشفها الهولندي Antonie van Leeuwenhoek في نهاية القرن 17. باحثو البيولوجيا ظلوا حتى القرن 19 يصنفون الكائنات الحية أحادية الخلية ضمن المملكتين الحيوانية والنباتية.
في القرن 19 ظهر المصطلحان التاليان:
- الحيوانات البدائية protozoans (مشتق من كلمة (zō̃ion) ζῷον التي تعني “حيوان”)
- النباتات البدائية protophytes (مشتق من كلمة (phytόn) φυτόν التي تعني “نبات”)
هذان المصطلحان يغطيان الكائنات الحية أحادية الخلية. “الحيوانات البدائية” protozoans هي تسمية أطلقت على الكائنات أحادية الخلية التي “تشبه الحيوانات” (من حيث امتلاكها القدرة على الحركةmotility ، ومن حيث اعتمادها على الكائنات الحية الأخرى في الغذاء heterotrophy). “النباتات البدائية” protophytes هي تسمية أطلقت على الكائنات أحادية الخلية التي “تشبه النباتات” (من حيث قدرتها على صنع غذائها بنفسها autotrophy عبر التركيب الضوئي photosynthesis).
في أواسط القرن 19 كثير من الباحثين صاروا يشعرون أن تقسيم الكائنات أحادية الخلية إلى حيوانات ونباتات هو تقسيم اعتباطي وغير علمي: هناك كائنات أحادية الخلية لا يمكن تصنيفها على أنها حيوان أو نبات. لهذا السبب اقترح الباحث الألمانيErnst Haeckel في عام 1866 إنشاء مملكة جديدة هي “مملكة الطلائعيات” Regnum Protista (المعنى الحرفي لكلمة protista هو “الأكثر بدائية”، ولكنني ترجمتها إلى “طلائعيات” لأن هذه هي الترجمة الشائعة). مملكة الطلائعيات كانت تضم “الكائنات المحايدة”، أي الكائنات التي ليست حيوانات ولا نباتات. لاحقا Haeckel عدّل تعريف هذه المملكة بحيث صارت تضم كل الكائنات وحيدة الخلية.
خلاصة تصنيفHaeckel هي ما يلي:
- مملكة الطلائعيات Regnum Protista (الكائنات وحيدة الخلية unicellular organisms)
- مملكة النباتات Regnum Plantae
- مملكة الحيوانات Regnum Animalia
في القرن العشرين ظهر اختراع مهم هو الميكروسكوب الإلكتروني electron microscope. هذا الميكروسكوب كشف أن الكائنات أحادية الخلية هي في الحقيقة صنفان مختلفان:
- كائنات بدائية النواة prokaryotes
- كائنات حقيقية النواة eukaryotes

الكائنات “حقيقية النواة” لها تركيب مشابه لتركيب الخلايا الحيوانية والنباتية، وأما الكائنات “بدائية النواة” فهي تتميز بتركيب أكثر بدائية. لهذا السبب الباحثون في أواسط القرن العشرين صاروا يرون أن الكائنات بدائية النواة هي الأقدم تطوريا. حسب النظرة الجديدة فإن الكائنات حقيقية النواة تطورت من الكائنات بدائية النواة، والكائنات متعددة الخلايا multicellular organisms (الحيوانات والنباتات) تطورت من الكائنات حقيقية النواة.
بناء ذلك ظهر تصنيف جديد يقسم الكائنات الحية إلى مجموعتين كبيرتين:
- إمبراطورية بدائيات النواة Imperium Prokaryota
- إمبراطورية حقيقيات النواة Imperium Eukaryota
الإمبراطورية الثانية تنقسم إلى ثلاث ممالك هي الطلائعيات والنباتات والحيوانات، وأما الإمبراطورية الأولى فهي أنتجت مملكة واحدة هي “مملكة الأحاديات” Regnum Monera.
مملكة النباتات حسب التصنيف التقليدي تضم الفطريات Fungi، ولكن هذا التصنيف هو تصنيف اعتباطي وليس تصنيفا علميا. Ernst Haeckel أدرك ذلك، ولهذا السبب هو أخرج الفطريات من مملكة النباتات وألحقها بمملكة الطلائعيات، ولكن معاصريه تجاهلوا ذلك وظلوا يصنفون الفطريات مع النباتات. في عام 1969 اقترح الباحث الأميركي Robert Whittaker إنشاء مملكة جديدة ضمن إمبراطورية حقيقيات النواة هي “مملكة الفطريات” Regnum Fungi. هذا الاقتراح لقي قبولا واسعا (على الأقل في أميركا) وبناء عليه صار تصنيف الكائنات الحية هو كما يلي:
- إمبراطورية بدائيات النواة Imperium Prokaryota
- مملكة الأحاديات Regnum Monera
- إمبراطورية حقيقيات النواة Imperium Eukaryota
- مملكة الطلائعيات Regnum Protista
- مملكة الفطريات Regnum Fungi
- مملكة النباتات Regnum Plantae
- مملكة الحيوانات Regnum Animalia
خلال العقود الأخيرة من القرن العشرين برز دور الدراسات الجزيئية والجينية. هذه الدراسات دمرت تماما التصنيفات التقليدية التي سردناها في الأعلى.
أول نتيجة للدراسات الجزيئية هي تقسيم الكائنات بدائية النواة prokaryotes إلى مجموعتين مستقلتين:
- البكتيريا Bacteria
- العتيقات Archaea

من غير المعروف أي المجموعتين هي الأقدم، ومن غير المعروف ما هي العلاقة بين هاتين المجموعتين وبين مجموعة حقيقيات النواة eukaryotes. الشيء الوحيد الذي يتفق عليه كثير من الباحثين حاليا هو أن الكائنات الحية تتوزع ضمن المجموعات الثلاث الظاهرة في الرسم التالي:

كل مجموعة من هذه المجموعات تسمى domain. المعنى الحرفي لهذه الكلمة هو “أرض سيّد” (land) belonging to a lord.
هناك نظرية ترى أن الأراضي الثلاث (bacteria و archaea و eukaryota) هي متحدرة من أصل مشترك كان موجودا في العصر السابق لظهور الخلايا.
الخلايا cells هي بنى معقدة، والباحثون لا يعتقدون أن الخلايا هي أقدم أشكال الحياة. هم يرون أن هناك أشكالا حية أكثر بدائية من الخلايا كانت موجودة قبل ظهور الخلايا، ولكن معلوماتهم عن هذه الأشكال الحية البدائية هي محدودة للغاية.
بالنسبة لمجموعة حقيقيات النواة eukaryotes فهناك حاليا جدل كبير حول تصنيف الكائنات المنضوية تحتها.
بعض الدراسات الحديثة تقسم حقيقيات النواة إلى مجموعتين كبيرتين:
- ثنائيات المجداف bikonts
- أحاديات المجداف unikonts
كلمة (kontós) κοντός تعني بالإنكليزية punting-pole (انظر القاموس). معنى punting-pole هو المجداف الذي يستخدم لتحريك قارب شبيه بالطوف يسمى punt (هذه صورته). إذن كلمتا bikonts و unikonts تعنيان حرفيا “ثنائيات المجداف” و”أحاديات المجداف”. المقصود بثنائيات المجداف هو الخلايا التي تملك سوطين two flagella، والمقصود بأحاديات المجداف هو الخلايا التي تملك سوطا واحدا one flagellum.
تصنيف ثنائيات المجداف يضم النباتات، وتصنيف أحاديات المجداف يضم الفطريات والحيوانات. بالنسبة للكائنات وحيدة الخلية فهي تتوزع بين التصنيفين.
بعض الدراسات ترى أن الكائنات ثنائية المجداف هي الأقدم تطوريا، وأن الكائنات أحادية المجداف هي مشتقة من كائن ثنائي المجداف.
كثير من الدراسات ترى أن الكائنات الأكثر بدائية بين ثنائيات المجداف (وبالتالي بين حقيقيات النواة) هي الكائنات المسماة excavates. هذا التصنيف يضم الكائن المسمى Euglena بالإضافة إلى عدد من الطفيليات المشهورة طبيا من قبيل Trypanosoma و Naegleria و Giardia و Trichomonas.

حسب بعض الدراسات فإن أقرب الكائنات إلى الـ excavates هي النباتات. هذا يوحي ربما بأن النباتات هي أكثر بدائية من الحيوانات والفطريات.
تصنيف النباتات حاليا هو محل جدل. بعض الدراسات تشطر المملكة التي كانت معروفة سابقا باسم مملكة النباتات إلى عدد كبير من المجموعات المستقلة، وبعض الدراسات تشطر مملكة النباتات إلى مجموعتين فقط:
- chromalveolates (“المجوّفات الصبغية”)
- archaeplastidans (“البلاستيديات العتيقة”)
المجموعة الثانية (archaeplastidans) تضم النباتات الحقيقية، وأما المجموعة الأولى فهي تضم أجناسا من الطحالب.
مجموعة الكائنات أحادية المجداف unikonts تقسم إلى مجموعتين:
- الحيوانات الأميبية amoebozoans
- خلفيات المجداف opisthokonts
مجموعة “الحيوانات الأميبية” amoebozoans هي جزء من المجموعة التي كانت تسمى سابقا “الأميبا” amoeba أو “شبيهات الأميبا” amoeboids. الجزء الآخر من مجموعة الأميبا يسمى rhizarians وهو ينتمي لمجموعة chromalveolates التي ذكرناها في الأعلى.

خلفيات المجداف opisthokonts هي المجموعة التي أنتجت الفطريات والحيوانات. بعض الدراسات تشطر هذه المجموعة إلى شطرين: شطر أنتج الفطريات وشطر آخر أنتج الحيوانات. الشطر الذي أنتج الحيوانات يسمى أحيانا holozoans (“الحيوانات الكاملة”). هذا الشطر أنتج مجموعة من الكائنات تسمى filozoans (“الحيوانات الخيطية”)، وهذه بدورها أنتجت مملكة الحيوانات التي تسمى أيضا metazoans (“الحيوانات التالية”).
أكثر الكائنات بدائية ضمن مملكة الحيوانات هي الإسفنجيات poriferans و”الحيوانات الصفيحية” placozoans. هذه الكائنات تصنف تحت عنوان “جانب الحيوانات” parazoans. بقية أعضاء مملكة الحيوانات تصنف تحت عنوان “الحيوانات التالية الحقيقية” eumetazoans.


الحيوانات التالية الحقيقية eumetazoans تنقسم إلى “الشعاعيات” radiates وثنائيات الجانب bilaterians.
الشعاعيات تضم المِشْطِيات ctenophores واللاسعات cnidarians. أجسام هذه الكائنات تتميز بالتناظر الشعاعي radial symmetry، بخلاف أجسام ثنائيات الجانب التي تتميز بالتناظر ثنائي الجانب bilateral symmetry. الفرق بين هذين النوعين من التناظر هو مذكور في هذا المقال.



مجموعة الحيوانات ثنائية الجانب bilaterians تضم عددا كبيرا جدا من الفروع. معظم هذه الفروع تدخل تحت مسمى الديدان worms (هذا المسمى هو مجرد تصنيف شعبي وليس تصنيفا علميا). إلى جانب الديدان مجموعة ثنائيات الجانب تضم أيضا مفصليات الأرجل arthropods وشوكيات الجلد echinoderms والحبليات chordates.
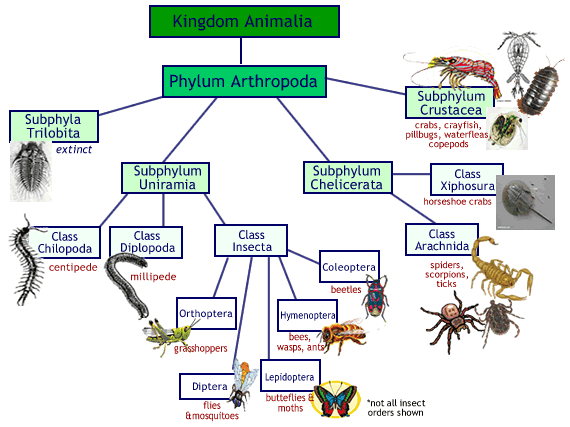

أبسط أنواع الحبليات chordates هي ربما “الحبليات الرأسية” cephalochordates المتمثلة بحيوان lancelet أو amphioxus (تحدثنا عنه في مقالات سابقة). الفقاريات vertebrates هي فرع من الحبليات. أنا تحدثت عن تطور الفقاريات في هذا المقال.

الحركة الانتقالية لدى الكائنات وحيدة الخلية
ما سبق هو عرض موجز لتطور الحيوانات. من هذا العرض يتبين أن الحيوانات هي فرع من حقيقيات النواة eukaryotes. اللافت هو أن خاصية الحركة motility هي على ما يبدو خاصية عتيقة جدا في حقيقيات النواة، بدليل أن الباحثين حاليا يصنفون حقيقيات النواة حسب عدد أسواطها إلى ثنائيات مجداف bikonts وأحاديات مجداف unikonts. حسب علمي فإن الوظيفة الأساسية للأسواط flagella هي الحركة الانتقالية locomotion. هذه الوظيفة تظهر واضحة لدى حقيقيات النواة البدائية المسماة excavates. هذه الكائنات تستخدم الأسواط للحركة الانتقالية.
بعض حقيقيات النواة تستخدم وسيلة أخرى للحركة الانتقالية هي الأهداب المتحركة motile cilia. الفرق بين عمل الأهداب المتحركة وبين عمل الأسواط هو طفيف.

هناك طريقة ثالثة للحركة الانتقالية لدى حقيقيات النواة هي “الأقدام الزائفة” pseudopodia التي تستخدمها الأميبا amoeba.

من الناحية التطورية الأسواطflagella هي ربما الأقدم. الأسواط هي موجودة لدى بدائيات النواة prokaryotes، ولكن الأهداب cilia والأقدام الزائفة pseudopodia هي غير موجودة لدى بدائيات النواة. أيضا لو نظرنا في مجموعة حقيقيات النواة فسنرى أن الـ excavates (التي هي الأكثر بدائية بين حقيقيات النواة) تعتمد على الأسواط وليس الأهداب أو الأقدام الزائفة.
بقايا الحركة السوطية والهدبية والقدمية الزائفة لدى البشر
أنماط الحركة التي سردناها في الأعلى ما زالت موجودة في أجسام البشر. بالنسبة للحركة عبر الأسواط فهي مثلا الطريقة التي يتحرك بواسطتها الحيوان المنوي sperm:

الحركة الهدبية هي موجودة لدى البشر في الخلايا ذات الأهداب المبطنة للقنوات التنفسية والتناسلية. حركة أهداب هذه الخلايا لا تؤدي لتحريك الخلايا ولكنها تؤدي لتحريك المفرزات الموجودة داخل القنوات التنفسية والتناسلية.

الحركة عبر الأقدام الزائفة هي الطريقة التي تتحرك بها خلايا الدم البيضاء:

الحركة الانتقالية لدى ثنائيات الجانب
بعض الحيوانات ثنائية الجانب ما زالت تعتمد على الأهداب المتحركة في حركتها الانتقالية. مثلا الديدان المسطحة platyhelminthes تعتمد على الأهداب المتحركة في حركتها الانتقالية، ولهذا السبب هي تتحرك ببطء شديد.
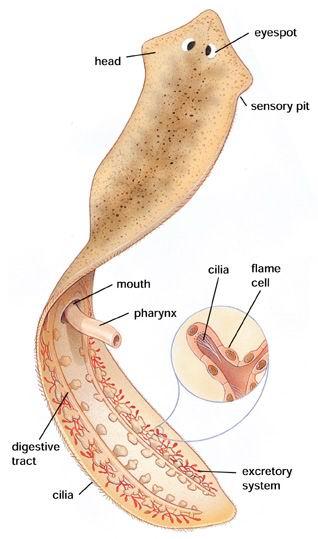
كثير من الديدان الأخرى تعتمد على نمط من الحركة يسمى التموّج undulation. هذا النمط من الحركة هو المشاهد لدى الديدان السهمية arrow worms والديدان المدورة roundworms وغيرها. هذا النمط من الحركة هو المشاهد أيضا لدى الحبليات والفقاريات البدائية.

تطور الحركة لدى ثنائيات الجانب هو مرتبط بظهور خاصية “التقطع المتتالي” metamerism في أجسامها.
بعض النظريات ترى أن التقطع المتتالي إنما ظهر بهدف تحسين الحركة. الجسم غير المقطع تقطيعا متتاليا هو عمليا أشبه بكتلة صخرية، لأن جوف الجسم coelom هو مملوء بالماء، والماء ينقل الضغط ويوزعه بشكل متساوي في جميع أنحاء الجسم. لو تقلصت إحدى عضلات جدار الجسم فستنتقل القوة الناتجة عن هذا التقلص عبر ماء الجوف نحو جميع مناطق الجسم. هذا الوضع يجعل من الصعب توجيه قوة التقلص العضلي نحو جزء محدد من الجسم، بالتالي الحيوان لا يمكنه أن يحرك جزءا محددا من جسمه ولكنه مضطر دائما للتحرك ككتلة واحدة.
الجسم المقطع إلى قطع متتالية metameres لا يعاني من هذه الصعوبة، لأن الحواجز التي تفصل بين القطع المتتالية تمنع انتقال الضغط عبر ماء الجوف من قطعة متتالية إلى أخرى. بالتالي الحيوان يمكنه أن يحرك كل قطعة متتالية على حدة. هذا يجعل جسم الحيوان مفصّلا hinged (بمعنى أنه يحوي مفاصل)، وبالتالي يحسن قدرة الحيوان على تحريك جسمه.
إذن التقطع المتتالي يعطي الحيوان القدرة على تنفيذ الحركات بدقة أكبر، ولكنه في نفس الوقت يخلق مشكلة جديدة هي ضرورة التنسيق الحركي motor coordination بين القطع المتتالية. تحريك قطعة واحدة لا يحتاج لتنسيق، ولكن تحريك عدد كبير من القطع هو بحاجة لتنسيق حتى لا تتعارض حركات القطع مع بعضها. تنسيق حركات القطع المتتالية يتم عبر الجهاز العصبي المركزي. هذا الجهاز يتكون لدى الديدان الحلقية annelids من حبلين بطنيين ventral cords وعقدتين دماغيتين cerebral ganglia. تكوين الجهاز العصبي المركزي لدى مفصليات الأرجل arthropods والحبليات chordates هو شبيه بتكوين الجهاز العصبي المركزي لدى الديدان الحلقية (كما بينا في مقالات سابقة).
الحركة التمعجية
معظم الديدان تعتمد على التموج undulation في حركتها الانتقالية، ولكن دودة الأرض earthworm (من الديدان الحلقية) طورت نوعا مميزا من الحركة الانتقالية يسمى peristalsis (“التَّمَعُّج”). المقطع التالي يبين هذه الحركة:
حركة “التمعج” هي موجودة لدى البشر ضمن القناة الهضمية، خاصة في المريء والأمعاء. حركة المريء والأمعاء لدى البشر تشبه حركة دودة الأرض الظاهرة في المقطع أعلاه، ولكنها ليست حركة انتقالية وإنما حركة تهدف لنقل الغذاء عبر القناة الهضمية.
الحركة التموجية لدى الفقاريات
الحركة التموجية undulation هي شائعة لدى ثنائيات الجانب البدائية. كثير من أنواع الديدان تتحرك بهذه الطريقة، وأيضا الحبليات والفقاريات البدائية تتحرك بهذه الطريقة.
المقصود بـ”الفقاريات البدائية” هو الأسماك، وخاصة الأسماك بلا فكين jawless fish كسمكة اللامبري lamprey. ما يلي مقطع يظهر حركة التموج لدى سمكة اللامبري:
حركة التموج الانتقالية هي موجودة أيضا لدى الأفاعي:
الأفاعي هي من الزواحف، ولكنها تعرضت لطفرة جينية جعلتها تفقد أطرافها limbs. لهذا السبب هي ارتدت نحو الحركة التموجية المميزة للفقاريات البدائية.
بقايا الحركة التموجية لدى الإنسان
في مقال سابق تحدثت عن بقايا الحركة التموجية لدى الإنسان. عضلات الجسم المسؤولة عن تنفيذ الحركة التموجية هي العضلات الباسطة المحورية axial extensor muscles. هذه العضلات هي مسؤولة عن بسط أو ثني العمود الفقري، وبالتالي هي مسؤولة عن الحركة التموجية. في الدماغ البشري هناك أنوية مسؤولة عن التحكم بهذه العضلات تقع في الدماغ الخلفي (hindbrain) rhombencephalon، وتحديدا في التشكل الشبكي reticular formation. هذه الأنوية هي مسؤولة لدى البشر عن عدد من المنعكسات البدائية التي تظهر خلال مرحلة الطفولة الباكرة. هذه المنعكسات ليس لها فائدة بالنسبة للبشر، ولهذا السبب يتم تثبيطها بشكل نهائي خلال مرحلة الطفولة، ولكن من الممكن أن هذه المنعكسات هي بقايا تطورية من زمن الفقاريات الباكرة، أي أن لها علاقة بالحركة الانتقالية التموجية.
الأنوية المسؤولة عن تقليص وإرخاء العضلات الباسطة المحورية (والتي كانت في السابق مسؤولة عن الحركة التموجية) اكتسبت لدى البشر وظيفة جديدة هي التحكم بالتوتر العضلي muscle tone للعضلات الباسطة المحورية. التوتر العضلي للعضلات الباسطة المحورية هو ضروري للمحافظة على استقرار الهيكل العظمي ومنع السقوط؛ لهذا السبب العضلات الباسطة المحورية تسمى لدى البشر باسم “العضلات المضادة للجاذبية” anti-gravity muscles.
الحركة بواسطة الأطراف الممفصلة
الحركة الانتقالية بواسطة الأطراف الممفصلة jointed limbs (الأطراف ذات المفاصل) هي شكل متطور من الحركة الانتقالية يميز نوعين من الحيوانات التي تعيش خارج الماء هما مفصليات الأرجل arthropods ورباعيات الأرجل tetrapods (رباعيات الأرجل هي فرع من الفقاريات يشمل الفقاريات التي تملك أربعة أطراف).
الحركة الانتقالية بواسطة الأطراف الممفصلة تشمل حركات المشي والركض والقفز والطيران بالأجنحة.
هذا الشكل من الحركة الانتقالية يتطلب وجود الأطراف الممفصلة ويتطلب القدرة على التحكم بها بشكل إيقاعي.
النواة الحمراء
أشرت في مقال سابق إلى النواة الحمراء red nucleus (باللاتينية: nucleus ruber). هذه النواة تظهر في مقطع الدماغ الأوسط المنقاري rostral midbrain عند مستوى التليلتين العلويتين superior colliculi:
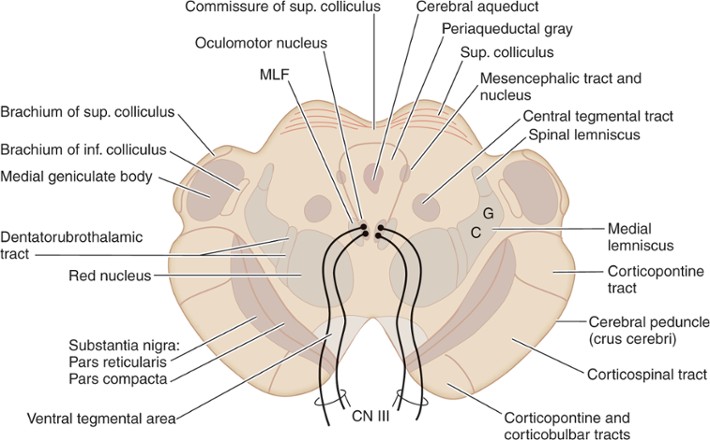

وظيفة النواة الحمراء مجهولة، ولكن هناك بعض النظريات حولها.
النواة الحمراء هي في الحقيقة نواتان اثنتان:
- النواة الحمراء كبيرة الخلايا magnocellular red nucleus (اختصارا RNm)
- النواة الحمراء صغيرة الخلايا parvocellular red nucleus (اختصارا RNp)
النواة الحمراء كبيرة الخلايا RNm تمثل القسم الذيلي من النواة الحمراء، والنواة الحمراء صغيرة الخلايا RNp تمثل القسم المنقاري من النواة الحمراء.
بعض الطروحات الحديثة ترى أن الوظيفة الأصلية للنواة الحمراء كبيرة الخلايا RNm هي تحريك عضلات الأطراف.
Although the RN can be differentiated in some fishes and limbless reptiles, a rubrospinal tract has so far been demonstrated only in rays which use their pectoral fins for locomotion [1]. The fins of the ray are likely precursors to the limbs of terrestrial vertebrates [7]. Rubrospinal tracts are also absent in boid snakes and sharks who use axial movements for locomotion rather than limbs [9]. Such data further implicates RNm and the corresponding rubrospinal tract in limb movement.
النواة الحمراء كبيرة الخلايا RNm هي مصدر لطريق عصبي نازل نحو الحبل الشوكي يسمى “الطريق الأحمر-الشوكي” rubrospinal tract. هذا الطريق هو غير موجود لدى “الفقاريات السفلى” (أي الأسماك)، باستثناء نوع من الأسماك الغضروفية يسمى rays (الاسم العلمي batoids). هذا النوع من الأسماك يتميز عن غيره بأنه يستخدم زعانفه للحركة الانتقالية locomotion.
الصور التالية تبين سمك الـ ray:


هناك اعتقاد بأن زعانف سمك الـ ray لها نفس أصل أطراف الفقاريات رباعية الأرجل.
الطريق الأحمر-الشوكي هو موجود لدى الزواحف باستثناء الأفاعي (التي تفتقد للأطراف).
الملاحظات السابقة تدعم الرأي القائل بأن وظيفة الطريق الأحمر-الشوكي هي تحريك الأطراف. هذا الطريق هو موجود حصرا لدى الفقاريات التي تتحرك بواسطة الأطراف.
النواة الحمراء صغيرة الخلايا RNp هي على ما يبدو غائبة لدى الأسماك والبرمائيات، وهي هامشية لدى بقية الفقاريات باستثناء الرئيسيات primates. اللافت هو أن نمو الـ RNp لدى الرئيسيات ترافق مع اضمحلال وضمور الـ RNm والطريق الأحمر-الشوكي النازل منها.
Studies of the mammalian red nucleus demonstrate that larger cells are found in RNm, whereas medium and small cells are typically found in RNp [3]. The volume of the nucleus taken up by these parts becomes reversed over the course of time and across species. In reptiles and birds the nucleus is composed almost entirely of large cells [4]. This large-celled RNm is the most likely source of the contralateral projections of the rubrospinal tract prominent in early vertebrates [1]. Yet as the rubrospinal tract has regressed in primates so has the histological presence of RNm. The number of large cells in the RN seems to diminish progressively as one passes from carnivores to baboons to anthropoids to man [2]. Morphological studies in humans show an increased development of the RNp – enough that RNm and RNp are completely independent of each other in humans [5].
[…]
The rubrospinal pathway and its cell bodies in RNm have progressively declined from lower mammals to primates [4]. Some researchers note that the RNm, initially involved in the automatic execution of learned motor skills, has been progressively replaced by the pyramidal system [1].
النواة الحمراء كبيرة الخلايا RNm اضمحلت بشكل تدريجي لدى الرئيسيات حتى وصلت إلى أقصى درجات اضمحلالها لدى البشر. الطريق الأحمر-الشوكي لدى البشر يصل فقط نحو القسم الرقبي cervical من الحبل الشوكي الذي يتحكم بالطرفين العلويين (الذراعان)، ولكنه لا يصل نحو القسم القَطَني lumbar من الحبل الشوكي الذي يتحكم بالطرفين السفليين (الرجلان). لدى بقية الثدييات (مثلا القطط) الطريق الأحمر-الشوكي يصل نحو القسم القطني من الحبل الشوكي.
في المقابل النواة الحمراء صغيرة الخلايا RNp نمت بشكل تدريجي لدى الرئيسيات حتى وصلت إلى أقصى درجات نموها لدى البشر. هذه النواة هي مصدر ألياف الطريق الأحمر-الزيتوني rubro-olivary tract الذي يصل نحو الزيتونة السفلية inferior olive.
الـ RNp تتلقى أليافا نازلة من قشرة المخ الحسية والمحركة والقشرة أمام الجبهية prefrontal (المسؤولة عن المعالجة المنطقية أو ما يسمى “الوظائف التنفيذية” executive functions). الـ RNp تتلقى أيضا أليافا من النواة المسننة في المخيخ dentate nucleus. اتصالات الـ RNp تشبه اتصالات قشرة المخ المحركة، ولهذا السبب بعض الباحثين يرون أن الـ RNp هي جزء من منظومة “الجهاز الهرمي” pyramidal system. هم يرون أن نمو الـ RNp ترافق مع نمو الجهاز الهرمي لدى الرئيسيات وهيمنة هذا الجهاز على “الجهاز خارج الهرمي” extrapyramidal system.
المقصود بالجهاز الهرمي هو المنظومة التحريكية التي تديرها قشرة المخ (المنظومة التحريكية الواعية)، والمقصود بالجهاز خارج الهرمي هو المنظومة التحريكية التي يديرها جذع الدماغ (المنظومة التحريكية غير الواعية). نحن عددنا طرق الجهاز الهرمي في مقالات سابقة (منها مثلا الطريق القشري-الشوكي corticospinal tract والطريق القشري-البصلي corticobulbar tract). أيضا نحن تحدثنا عن بعض طرق الجهاز خارج الهرمي في مقالات سابقة (منها الطريق الشبكي-الشوكي reticulospinal tract والطريق الدهليزي-الشوكي vestibulospinal tract).
النواة الحمراء كبيرة الخلايا RNm تستقبل أليافا نازلة من قشرة المخ المحركة، وتستقبل أليافا صاعدة من الحبل الشوكي تسمى الطريق الشوكي-الأحمر spinorubral tract. هذا الطريق يشبه الطريق الشوكي-الشبكي spinoreticular tract والطريق الشوكي-الدهليزي spinovestibular tract. اتصالات الـ RNm تتناسب مع كونها جزءا من الجهاز خارج الهرمي extrapyramidal system.
الـ RNm تتلقى أليافا من نواة مخيخية تسمى “النواة المتوسطة” interposed nucleus. على الأغلب أن وظيفة هذه النواة لها علاقة بتحريك الأطراف. النواة المسننة dentate nucleus (التي ترسل أليافا نحو الـ RNp ونحو قشرة المخ المحركة) هي مرتبطة بالجهاز الهرمي. هناك نواة مخيخية ثالثة هي “النواة الأَوْجِية” fastigial nucleus. هذه النواة هي أقدم الأنوية المخيخية وهي ترسل أليافا نحو الأنوية الدهليزية. على الأغلب أن وظيفة النواة الأوجية لها علاقة بتحريك العضلات الباسطة المحورية للجسم (تشرف على تنظيم وضعية الجسم posture عبر التحكم بالتوتر العضلي muscle tone).
الوضعية مزالة القشرة
هناك حالة طبية تسمى “الوضعية مزالة الدماغ” decerebrate posture. هذه الوضعية تظهر لدى الأشخاص المصابين بأذية كبيرة في جذع الدماغ تحت مستوى النواة الحمراء.

في هذه الوضعية هناك تقلص عام للعضلات الباسطة المحورية (وهو ما يؤدي إلى انبساط أو انتصاب الجسم).
ما هو سبب هذه الوضعية؟
السبب هو انقطاع الألياف النازلة من قشرة المخ نحو التشكل الشبكي والأنوية الدهليزية. هذه الألياف تمارس تأثيرا مثبطا أو منظما للأنوية الشبكية والدهليزية المسؤولة عن تقليص العضلات الباسطة المحورية (عبر الطريق الشبكي-الشوكي reticulospinal tract والطريق الدهليزي-الشوكي vestibulospinal tract).
هناك وضعية أخرى ملفتة أكثر من الوضعية سالفة الذكر تسمى “الوضعية مزالة القشرة” decorticate posture. هذه الوضعية تظهر لدى الأشخاص المصابين بأذية فوق مستوى النواة الحمراء.

هذه الوضعية تشبه وضعية المومياء المصرية. في هذه الوضعية هناك انبساط في عضلات الجسم ما عدا عضلات الطرفيين العلويين التي هي في حالة انقباض أو ثني.
تفسير هذه الوضعية هو أن الأذية فوق مستوى النواة الحمراء تزيل التثبيط القشري عن النواة الحمراء كبيرة الخلايا RNm، وهذا يؤدي إلى تنشيط الطريق الأحمر-الشوكي rubrospinal tract الذي يؤثر في الطرفين العلويين دون الطرفين السفليين (لأن أليافه لدى البشر لا تنزل نحو القسم القطني lumbar من الحبل الشوكي).
الوضعية مزالة القشرةdecorticate توحي بأن الطريق الأحمر-الشوكي يمارس تأثيرا مقبضا لعضلات الأطراف. هذا التأثير يهدف في الأصل لتحريك الأطراف.
وظيفة النواة الحمراء كبيرة الخلايا
في الأعلى قلنا أن وظيفة النواة الحمراء كبيرة الخلايا RNm هي على الأغلب تحريك الأطراف بهدف الحركة الانتقالية locomotion. نشوء هذه النواة هو مرتبط بظهور الحركة الانتقالية عبر تحريك الأطراف.
السؤال هو لماذا ضمرت هذه النواة لدى الرئيسيات عموما والبشر خصوصا؟
الميزة الأساسية التي تميز الرئيسيات عن بقية الحيوانات هي المهارة اليدوية. الطرف limb لدى الرئيسيات لم يعد مجرد أداة للحركة الانتقالية على غرار السوط flagellum أو الهدب المتحرك motile cilium أو القدم الزائفة pseudopodium، ولكنه تحول تدريجيا إلى عضو للإمساك بالأشياء وتفحصها والتأثير فيها.
هذا التغير في وظيفة الأطراف يظهر بشكل جلي لدى البشر. الطرفان العلويان لدى البشر لم يعد لهما علاقة تذكر بتحريك الجسم، أي أن وظيفتهما الأصلية تلاشت تماما.
بالنسبة للطرفين السفليين فهما ما زالا يؤديان وظيفة تحريك الجسم، ولكن الطريقة التي يؤديان بها هذه الوظيفة تغيرت كثيرا. الطريقة التي يمشي بها البشر لا تكاد تشبه الطريقة التي تمشي بها غالبية الثدييات. البشر يعتمدون المشية ثنائية القدم bipedal وليس المشية رباعية القدم quadrupedal التي تميز غالبية رباعيات الأرجل tetrapods. هناك اختلافات كبيرة بين المشيتين. مثلا البشر لا يكادون يثنون أرجلهم عند المشي، وحتى عند الوقوف أو النوم هم في الغالب لا يثنون أرجلهم. في الحقيقة البشر يبقون أرجلهم منبسطة في أغلب الأوقات. هذا يختلف عن الحال لدى الحيوانات ذات المشية رباعية الأرجل. تلك الحيوانات تبقي أرجلها مثنية في أغلب الأوقات. بما أن وظيفة الـ RNm هي ثني الأطراف فليس مستغربا أن هذه النواة فقدت تأثيرها على الطرفيين السفليين لدى البشر. تأثير هذه النواة على الطرفيين السفليين لم يعد مفيدا للبشر.
إذن النواة الحمراء كبيرة الخلايا RNm أصبحت قليلة الفائدة للبشر، لأن البشر ما عادوا يثنون أطرافهم الخلفية إلا فيما ندر. هم ما زالوا يثنون أطرافهم الأمامية، ولكنهم لا يمشون على هذه الأطراف. هذه الأطراف صارت تستخدم في الأساس للإمساك بالأشياء والتأثير فيها. وظيفة الإمساك بالأشياء والتأثير فيها تتم عبر أصابع اليدين. التحكم بأصابع اليدين يتم عبر الجهاز الدهليزي (قشرة المخ). لهذا السبب الـ RNm لم تعد عضوا مهما ولكنها صارت عضوا شبه أثري vestigial.
صفحة ويكيبيديا تقول أن النواة الحمراء تتحكم بحركة الزحف crawling لدى الأطفال الرضع، وأنها مسؤولة عن أرجحة الذراعين أثناء المشي، ولكن مصدر هذه المعلومات غير مذكور.
وظيفة النواة الحمراء صغيرة الخلايا
وظيفة النواة الحمراء صغيرة الخلايا RNp هي أقل وضوحا من وظيفة نظيرتها كبيرة الخلايا RNm. كما قلنا في الأعلى فإن اتصالات هذه النواة أوحت للباحثين أنها جزء من الجهاز الهرمي. الجهاز الهرمي هو مسؤول عن الحركات المعقدة التي تميز البشر من قبيل تحريك الأصابع والكلام. هذه الدراسة مثلا تربط بين نمو الـ RNp لدى البشر وبين تطور المهارات اليدوية والكلامية لديهم.
حسب ما فهمت فإن الـ RNp لا ترسل أية ألياف نازلة نحو الحبل الشوكي. هذه النواة هي في الأصل نواة حسية (صغيرة الخلايا parvocellular) وليست نواة تحريكية.
في الحقيقة اتصالات الـ RNp هي شبيهة باتصالات النواة الزيتونية السفلية inferior olivary nucleus، وهناك حتى اتصال مباشر بين النواتين عبر الطريق الأحمر-الزيتوني rubro-olivary tract. النواة الزيتونية السفلية (كالـ RNp) تتلقى أليافا من نواة المخيخ المسننة dentate nucleus.
بناء على التشابه في الأصل التكويني والاتصالات العصبية بين الـ RNp والنواة الزيتونية السفلية أنا أشعر بأن هاتين النواتين هما مكملتان لبعضهما ومن الممكن أنهما تؤديان نفس الوظيفة (تحديث معلومات المخيخ). النواة الزيتونية السفلية تستقبل بيانات قادمة من الحبل الشوكي، والـ RNp تستقبل بيانات قادمة من قشرة المخ. التجميع النهائي لهذه البيانات يتم في النواة الزيتونية السفلية، ومن هناك يتم إرسال إشارات التحديث أو التصحيح نحو المخيخ (والله أعلم).
في المقال القادم سأكمل الحديث في موضوع الحركة الانتقالية بواسطة الأطراف.
