في المقالين الأخيرين من هذه السلسلة بدأت الحديث عن جذع الدماغ brainstem وعن تركيبه ووظائفه.
في أحد هذين المقالين تحدثت عن توزع المادتين الرمادية والبيضاء في مقطع النخاع المستطيل الذيلي (السفلي) caudal (inferior) medulla oblongata. قبل أن أكمل الحديث عن وظائف جذع الدماغ سأتحدث قليلا عن توزع المادتين الرمادية والبيضاء في بقية مقاطع جذع الدماغ.
النخاع المستطيل المنقاري
المقصود بالنخاع المستطيل الذيلي caudal medulla oblongata هو القسم السفلي من النخاع المستطيل الذي يقع أعلى الحبل الشوكي spinal cord. المقصود بالنخاع المستطيل المنقاري rostral medulla oblongata هو القسم العلوي من النخاع المستطيل الذي يقع أسفل الجسر pons.
الفرق الأساسي بين النخاعين الذيلي والمنقاري هو وجود المخيخ cerebellum على ظهر النخاع المنقاري. المخيخ موجود أيضا على ظهر الجسر.

بين المخيخ وجذع الدماغ يوجد تجويف مملوء بالسائل الدماغي الشوكي cerebrospinal fluid. هذا التجويف يسمى “البطين الرابع” fourth ventricle. هذا التجويف هو عبارة عن توسع في القناة المركزية central canal الموجودة في وسط الحبل الشوكي والنخاع الذيلي.

الصورة التالية هي لمقطع في النخاع المستطيل الذيلي:
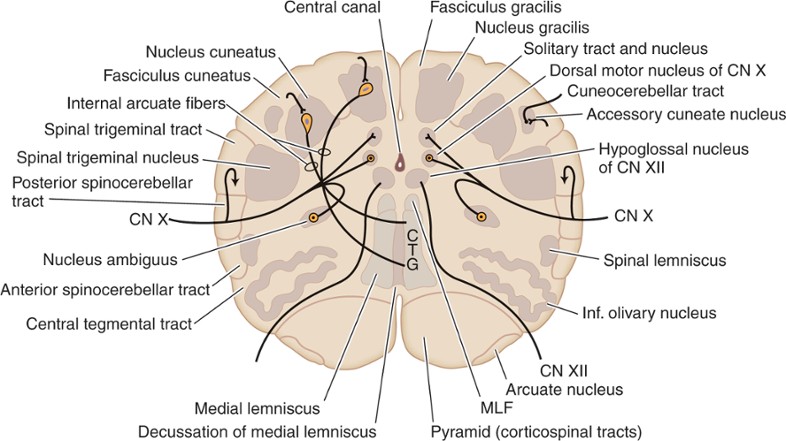
هذا المقطع تحدثنا عنه سابقا. لاحظ في هذا المقطع وجود القناة المركزية central canal كما في مقطع الحبل الشوكي.
الصورة التالية هي لمقطع في النخاع المستطيل المنقاري:
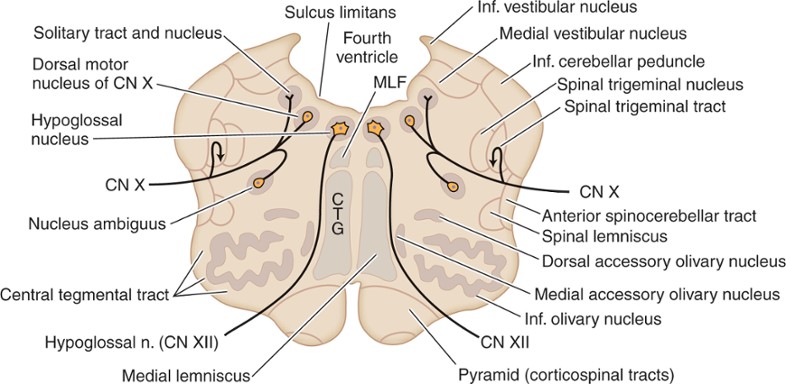
لاحظ في هذا المقطع أن القناة المركزية انفتحت من الجهة الظهرية وكونت تجويفا كبيرا. هذا التجويف هو قعر البطين الرابع fourth ventricle. الأنوية الرمادية التي كانت تحيط بالقناة المركزية في المقطع السابق صارت تقع أسفل قعر البطين الرابع، وهذا أدى إلى تغيير التوزع الوظيفي: الأنوية المحركة الجسمانية somatic motor (مثلا نواة العصب القحفي تحت اللساني hypoglossal nucleus) صارت تقع في الجهة الداخلية أو الإنسية medial للأنوية الرمادية (بعدما كانت تقع في الجهة البطنية ventral)، والأنوية التأثيرية الحشوية visceral effector (مثلا النواة الظهرية للعصب القحفي dorsal nucleus of vagus nerve) صارت تقع على الجهة الخارجية أو الوحشية lateral للأنوية المحركة الجسمانية (بعدما كانت تقع على جهتها الظهرية dorsal). الأنوية الحسية الحشوية visceral sensory (مثلا نواة الطريق المفرد nucleus of the solitary tract) والأنوية الحسية الجسمانية somatic sensory صارت تقع في الجهة الخارجية أو الوحشية lateral للأنوية الرمادية (بعدما كانت تقع في الجهة الظهرية dorsal).
الشكل التالي يوضح التغير في مواقع الأنوية الرمادية:

النواتان الحسيتان الجسمانيتان المسماتان “النواة النحيلة” nucleus gracilis و”النواة وتدية الشكل” nucleus cuneatus لم تعودا موجودتين في مقطع النخاع المستطيل المنقاري. بدلا منهما ظهرت أنوية تحمل مسمى الأنوية الدهليزية vestibular nuclei (في المقطع هناك نواتان دهليزيتان ظاهرتان، ولكنهما ليستا ملونتين باللون الرمادي ولا أدري لماذا).
لو صعدنا أكثر نحو منطقة اتصال النخاع المستطيل مع الجسر فستظهر أيضا النواتان القوقعيتان cochlear nuclei على الجانب الوحشي (الخارجي) للأنوية الدهليزية. المقطع التالي يبين هذه الأنوية.

في هذا المقطع تظهر نواتان دهليزيتان. العدد الكلي للأنوية الدهليزية هو أربع (إنسية ووحشية وسفلية وعلوية). العدد الكلي للأنوية القوقعية هو اثنتان (ظهرية وبطنية). الأنوية الدهليزية تستقبل ألياف حس الاتزان balance القادمة من الأذن الداخلية inner ear عبر العصب الدهليزي vestibular nerve، والنواتان القوقعيتان تستقبلان ألياف حس السمع القادمة من الأذن الداخلية عبر العصب القوقعي cochlear nerve. مجموع العصبين الدهليزي والقوقعي يسمى العصب الدهليزي القوقعي vestibulocochlear nerve (العصب القحفي الثامن).


وظيفة القوقعة هي استشعار اهتزازت الهواء التي تصل إلى طبلة الأذن على شكل موجة هوائية ثم تنتقل عبر عظام الأذن الوسطى إلى الأذن الداخلية. القوقعة تميز بين اهتزازات الهواء المختلفة حسب ترددها frequency، وهذا هو ما يعطي البشر القدرة على التمييز بين الأصوات “الحادة” (عالية التردد) والأصوات “الغليظة” (منخفضة التردد).
وظيفة الدهليز هي استشعار التسارع الخطي linear acceleration، أي أنه يستشعر حركة الرأس بشكل مستقيم في أي اتجاه. وظيفة الأقنية نصف الدائرية هي استشعار التسارع الزاوي angular acceleration، أي أنها تستشعر حركة الرأس بشكل دائري في أي اتجاه. إذن وظيفة الدهليز والأقنية نصف الدائرية هي استشعار حركة الرأس.

حس السمع وحس الاتزان هما من “الأحاسيس الخاصة” special senses. عبارة “الأحاسيس الخاصة” يستخدمها الباحثون لوصف الأحاسيس التي توجد ضمن أعضاء معينة وليس في جميع أنحاء الجسم. الأحاسيس الخاصة عند الفقاريات توجد في الرأس دون بقية الجسم (الشم والبصر والسمع والاتزان والذوق). الأحاسيس التي توجد في جميع أنحاء الجسم تسمى “الأحاسيس العامة” general senses (مثلا حس الألم والحرارة والاهتزاز والحس الذاتي proprioception إلخ).
الألياف العصبية التي تنقل أحاسيس البصر والسمع والاتزان تعتبر أليافا جسمانية somatic، والألياف التي تنقل حسي الشم والذوق تعتبر أليافا حشوية visceral.
على ما أظن فإن الأحاسيس الخاصة كانت في الأصل أحاسيس عامة، ولكن في زمن ظهور الحبليات chordates (أو قبله) تركزت بعض الأحاسيس العامة في الرأس. هذه القضية هي فرع من قضية “الرأسنة” cephalization التي أدت لنشوء الدماغ.
الأحاسيس الخاصة عند الحشرات يمكن أن توجد في أي مكان من الجسم وليس فقط في الرأس. مثلا مستقبلات حس الذوق لدى بعض الحشرات (الفراش والذباب) هي موجودة على الأقدام. أيضا مستقبلات حس السمع (التي تسمى “الأعضاء الطبلية” tympanal organs) هي موجودة على الصدر أو البطن لدى الكثير من الحشرات. بالنسبة لحسي الشم والبصر عند الحشرات فهما موجودان في الرأس، ولكن ترتيبهما مختلف عن الترتيب المشاهد لدى الفقاريات (كما بينت في مقال سابق)، وحتى طبيعتهما هي طبيعة مختلفة (العين لدى الفقاريات هي مشتقة من الدماغ البيني diencephalon، ولكن العين المركبة لدى الحشرات هي ليست مشتقة من الدماغ أو حتى من النسيج العصبي). أنا لا أرى تشابها في توزع الأحاسيس الخاصة لدى الحشرات ونظيرتها لدى الفقاريات. ما يبدو لي هو أن عملية “التخصص الحسي” حصلت بشكل مستقل لدى كل من الفقاريات ومفصليات الأرجل. الأحاسيس الخاصة لدى الفقاريات ومفصليات الأرجل لا تبدو موروثة من أصل مشترك.
حس السمع لدى البشر يستخدم أساسا في التواصل communication مع الكائنات الأخرى، ولكن الوظيفة الأصلية لهذا الحس لم تكن التواصل بقدر ما أنها كانت المساعدة على الإحساس بمواقع الأجسام المتحركة المحيطة بالجسم. هذه الوظيفة الأصلية لحس السمع تظهر بوضوح لدى الوطواط الذي يعتمد على حس السمع بشكل كبير لتحديد مواقع الأجسام المحيطة به.
الأسماك تعتمد بشكل كبير على حس السمع لتحديد مواقع الأجسام المتحركة المحيطة بها، ولكن اللافت لدى الأسماك هو أن حس السمع لديها ليس مقصورا على الأذن ولكنه موجود أيضا ضمن عضو يسمى “الخط الجانبي” lateral line.

وظيفة الخط الجانبي لدى الأسماك هي تقريبا نفس وظيفة الأذن، أي تحديد مواقع الأجسام المتحركة المحيطة بالسمكة. الباحثون يعتقدون أن الخط الجانبي والأذن لهما أصل واحد.
حس البصر يساعد أيضا على تحديد مواقع الأجسام المحيطة، ولكن حس البصر له نواقص كثيرة. أولا هذا الحس لا يغطي جميع الاتجاهات، بخلاف حس السمع الذي يغطي كل الاتجاهات المحيطة بالكائن (مثلا لو أتى شخص من خلفي فإنني لن أراه ولكنني سأسمع صوته). حس البصر يعتمد على الضوء، وبالتالي هو غير مفيد في الظلام. حس السمع لا يتأثر بغياب الضوء وهو يعمل في الظلام.
إذن حس السمع له مزايا غير موجودة في حس البصر.
حسب ما أعلم فإن حس البصر هو أقدم من حس السمع. حس البصر موجود بشكل بدائي لدى بعض الكائنات وحيدة الخلية protists (مثلا لدى الكائنات المسماة euglena)، وهو موجود لدى بعض اللاسعات cnidarians (مثلا لدى قنديل البحر jellyfish). بالنسبة لحس السمع فلا أدري متى كانت بداية ظهوره، ولكنه على ما يبدو خاص بثنائيات الجانب bilaterians (مثلا مفصليات الأرجل والفقاريات). أكثر ثنائيات الجانب بدائية هي الديدان، والديدان حسب علمي لا تملك حس السمع (ولكن ديدان الأرض earthworms يمكنها الشعور بالاهتزاز).
حس الاتزان هو أقدم من ثنائيات الجانب بدليل وجوده لدى قنديل البحر jellyfish. حس الاتزان لدى الحيوانات غير الفقارية يعتمد على مستقبلات حسية تسمى “أكياس الاتزان” statocysts. لدى الفقاريات هناك نوع آخر من مستقبلات الاتزان هو “الأحجار الأذنية” otoliths. العلاقة بين “أكياس الاتزان” statocysts و”الأحجار الأذنية” otoliths هي غير واضحة ولكن غالبية الباحثين لا يعتقدون بوجود أصل مشترك بينها. هذه المستقبلات هي متشابهة في الوظيفة وآلية العمل، ولكنها نشأت بشكل مستقل لدى كل من الفقاريات والحيوانات غير الفقارية.
الأحجار الأذنية otoliths تعتمد على نوع من الخلايا يسمى الخلايا الشعرية hair cells. مستقبلات السمع في القوقعة تعتمد أيضا على الخلايا الشعرية، والخط الجانبي لدى الأسماك يعتمد على الخلايا الشعرية. إذن هذه الخلايا هي أساس حس التوازن والسمع لدى الفقاريات. أصل هذه الخلايا هو غير واضح، ولكن إحدى النظريات الحديثة ترى أن هذه الخلايا كانت في السابق تغطي الجسم ولاحقا تخصصت في الأذن الداخلية والخط الجانبي:
http://onlinelibrary.wiley.com/doi/10.1002/ar.22573/pdf
The difference is that this theory does not presume the prior evolution of lateral line organs, and in fact suggests that the hair cell and its precursor the ciliated mechanosensory cell were present well before the lateral line and inner ear as organ systems evolved. This hypothesis also does not presuppose that hair cells evolved within a statocyst organ, but rather as diffuse cells in the skin, which later became aggregated to evolve into the ear.
Jeremy S. Duncan and Bernd Fritzsch, Evolution of Sound and Balance Perception: Innovations That Aggregate Single Hair Cells Into the Ear and Transform a Gravistatic Sensor Into the Organ of Corti
ما سبق يعني أن حس السمع هو مشتق من حس الاتزان. هذا الكلام ليس غريبا لأن وظيفة حس السمع تشبه وظيفة حس الاتزان. الوظيفة الأصلية لحس الاتزان هي منح الكائن القدرة على الإحساس بحركة جسمه. الوظيفة الأصلية لحس السمع هي منح الكائن القدرة على الإحساس بحركة الأجسام المحيطة به. إذن الفرق بين حسي الاتزان والسمع هو ليس في وظيفة الحس ولكن في اتجاهه: حس الاتزان موجه نحو الذات، وحس السمع موجه نحو الخارج.
هذا الفرق يشبه الفرق بين المستقبلات الكيماوية chemoreceptors التي تستشعر تغيرات تراكيز المواد الكيماوية داخل الجسم وبين مستقبلات حسي الشم والذوق التي تستشعر تغيرات تراكيز المواد الكيماوية خارج الجسم.
حسا الشم والذوق يشبهان بعضهما. كلاهما يعتمدان على مستقبلات كيماوية موجهة نحو المحيط الخارجي، ولكن الفرق بينهما هو أن مستقبلات الشم تتحسس المواد الكيماوية الغازية، ومستقبلات الذوق تتحسس المواد الكيماوية السائلة. عند الحيوانات التي تعيش في الماء لا يوجد فرق عملي بين هذين الحسين.
هناك أيضا شبه بين حس الاتزان/السمع وبين حس الاهتزاز وبقية الأحاسيس الميكانيكية الموجودة في الجلد (الضغط والشد إلخ). آلية عمل الخلايا الشعرية hair cells تشبه آلية عمل المستقبلات الميكانيكية mechanoreceptors الموجودة في الجلد.
على ما أظن فإن جميع الأحاسيس الحيوانية يمكن إرجاعها إلى الأحاسيس التالية:
- الحس الكيماوي chemoreception
- الحس الضوئي photoreception
- الحس الميكانيكي mechanoreception
- الحس الحراري thermoreception
- حس الألم nociception
مستقبلات هذه الأحاسيس كانت في الأصل مستقبلات عامة تغطي الجسم، ولكن لاحقا حصلت عملية تخصص أدت إلى حصر الحس الضوئي والحس الكيماوي في مناطق محددة من الجسم. الحس الميكانيكي تخصص بشكل جزئي: بعض أنواع الحس الميكانيكي (المعتمدة على الخلايا الشعرية) تخصصت وانحصرت في منطقة الرأس (تحديدا في الأذن).
لو عدنا إلى مقطع النخاع المستطيل المنقاري فسنرى أن الأنوية الدهليزية والقوقعية تقع في نفس منطقة النواتين النحيلة ووتدية الشكل. هذا ليس مفاجئا لأن حسي الاتزان والسمع هما كما قلنا مشتقان من الحس الميكانيكي العام. النواتان النحيلة ووتدية الشكل تستقبلان حس الاهتزاز وحس الضغط والحس الذاتي proprioception (الحس الذاتي يعني حس الشد في العضلات وأوتارها) من العمودين الظهريين dorsal columns. هذه الأحاسيس الميكانيكية “العامة” هي أقرب الأحاسيس إلى حسي الاتزان والسمع (الحسان الميكانيكيان “الخاصان”).
