نزار نيوف هو ناقد لاذع، وهو لا يترك أمرا لا يعجبه دون أن يبدي رأيه النقدي اللاذع فيه.
ولكن في الآونة الأخيرة لاحظت أنه ينتقد بشدة حياة الرعي والرعاة وما يتصل بها.
هو غضب جدا من باسل ديوب لأنه عيّر الناس بمهنهم وأصولهم الاجتماعية، ولكنه في نفس الوقت يعيّر أحمد الجربا بكونه راعيا للغنم، وهو يتهم بشار الأسد بأنه راع للغنم على سبيل الذم:
قال نيوف “قبل حوالي سبع سنوات ، طلب هيثم سطايحي من شيخ الشبيحة (الأسد)، وكان رئيس مكتبه آنذاك، أن يلغي قراراته الثلاثة بطرد إخوتي الثلاثة من وظائفهم ، بعد أن لفت انتباهه إلى الأمر وطلب منه ذلك أحد الأساتذة في جامعة تشرين في اللاذقية ، وإلى أن السلطة انتقمت من أبي وأخوتي الثلاثة بسببي دون أن يكون لهم علاقة بأنشطتي. إلا أن شيخ الشبيحة أجاب سطايحي بالحرف الواحد: كيف بدي رجعهم لوظائفهم وأخوهم ما عليه شغل غير الحكي عليي؟ خلوه يوقف حكي عليي، بيرجعوا أخوانه وأبوه لشغلهم؟”!!؟ وقال نيوف”جواب من هذا النوع لا يمكن أن يقوله إلا راعي غنم أو ماعز مثل أحمد الجربا، مع احترامنا وتقديرنا للرعيان الحقيقيين جميعا”.
لو قرر النظام السوري توزيع سيارات على أسر الشهداء فهل كان نيوف سيسخر من ذلك؟
ما الذي يعيب الماعز؟
السخرية من الرعاة هي بالمناسبة ليست خصلة خاصة بنيوف، بل هي طبع عام موجود في بعض المناطق السورية. في الثقافة الحلبية مهنة الرعي هي مهنة مذمومة ومكروهة.
هذا الكره للرعاة هو شيء قديم كان موجودا منذ الألفية الثانية قبل الميلاد. أنا في السابق تحدثت عن قصة قايين وهابيل التوراتية وكيف أن هذه القصة في الأصل ترمز للصراع بين الفلاحين والرعاة في الألفية الثانية قبل الميلاد.
في رأيي أن السخرية من الرعاة لمجرد كونهم رعاة هو أمر عنصري ومعيب.
أنا ما كنت لأتحدث عن هذا الموضوع لولا أنني قرأت قبل أيام كلاما لنزار نيوف يهاجم فيه بشدة باسل ديوب لأنه عير الناس بأصولهم الاجتماعية.
ذكرنا في المقال السابق أن الجهاز العصبي المركزي لدى ثنائيات الجانب bilaterians الأولى كان عبارة عن حبل ممتد على طول الجسم يسمى الحبل العصبي nerve cord.
الحبل العصبي عند الحبليات chordates يمتد على طول الظهر بمحاذاة الحبل الظهري notochord (ولاحقا بمحاذاة العمود الفقري لدى الفقاريات). عند ثنائيات الجانب غير الحبلية (الديدان worms ومفصليات الأرجل arthropods) الحبل الظهري يمتد على طول البطن وليس الظهر.
في الحقيقة الديدان ومفصليات الأرجل تملك حبلين عصبيين وليس حبلا واحدا:
الجهاز العصبي لدودة مسطحة platyhelminthالجهاز العصبي لدودة حلقية annelid (دودة الأرض earthworm)
الديدان هي الحيوانات الأكثر بدائية ضمن مجموعة “ثنائيات الجانب” bilaterians، بمعنى أنها ربما أكثر الحيوانات المعاصرة شبها بثنائيات الجانب الباكرة. ثنائيات الجانب سميت بهذا الاسم لأن أجسامها تتكون من شطرين متناظرين حول المستوي السهمي sagittal plane (المستوي الذي يمر من السطح البطني للجسم إلى السطح الظهري قاسما الجسم إلى شطرين أيمن وأيسر). لو قسمنا حيوانا ثنائي الجانب إلى شطرين أيمن وأيسر فإن الشطرين يجب أن يكونا متناظرين:
على اليمين حيوان ثنائي الجانب (من مفصليات الأرجل) وعلى اليسار كائن ذو تناظر شعاعي radial symmetry
الكائنات ذات التناظر الشعاعي radial symmetry يمكن أن نقسمها إلى شطرين متناظرين مهما كان اتجاه المستوي الرأسي-الذيلي الذي سيتم الشطر عبره. التناظر الشعاعي هو شائع لدى الحيوانات البدائية (الإسفنجيات والمِشْطِيات واللاسعات)، وأيضا موجود بكثرة في النباتات.
وجود حبلين عصبيين في الجسم (حبل أيمن وأيسر) يتوافق مع مبدأ التناظر ثنائي الجانب، وبالتالي المفترض هو أن الكائنات ثنائية الجانب الباكرة كانت تملك حبليين عصبيين كالديدان ومفصليات الأرجل (“مفصليات الأرجل” arthropods هو تصنيف يشمل الحشرات وأم أربعة وأربعين والعناكب والسرطانات).
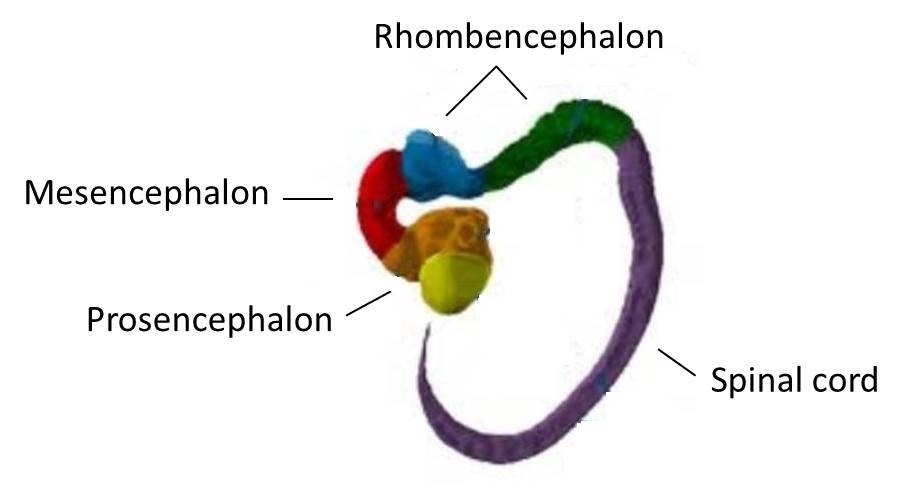
تصوران افتراضيان لأول حيوان ثنائي الجانب urbilaterian
تطور الدماغ
الحيوانات ثنائية الجانب تتميز بأن لها رأسا يحوي كتلة عصبية (دماغ). نشوء الرأس الذي يحوي الدماغ يسمى cephalization.
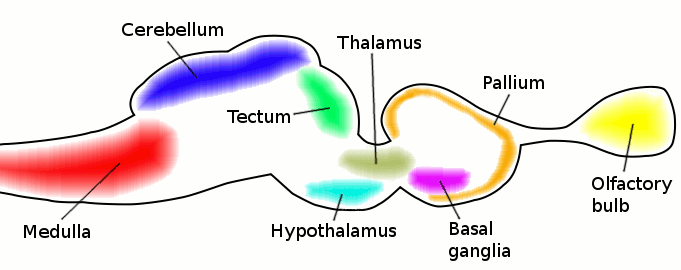
نشوء الدماغ عند الحبليات الباكرة بدأ بانتفاخ في الحبل العصبي في منطقة الرأس. عند الفقاريات الباكرة الدماغ كان مكونا من ثلاثة حويصلات على نحو شبيه بهذا الشكل:
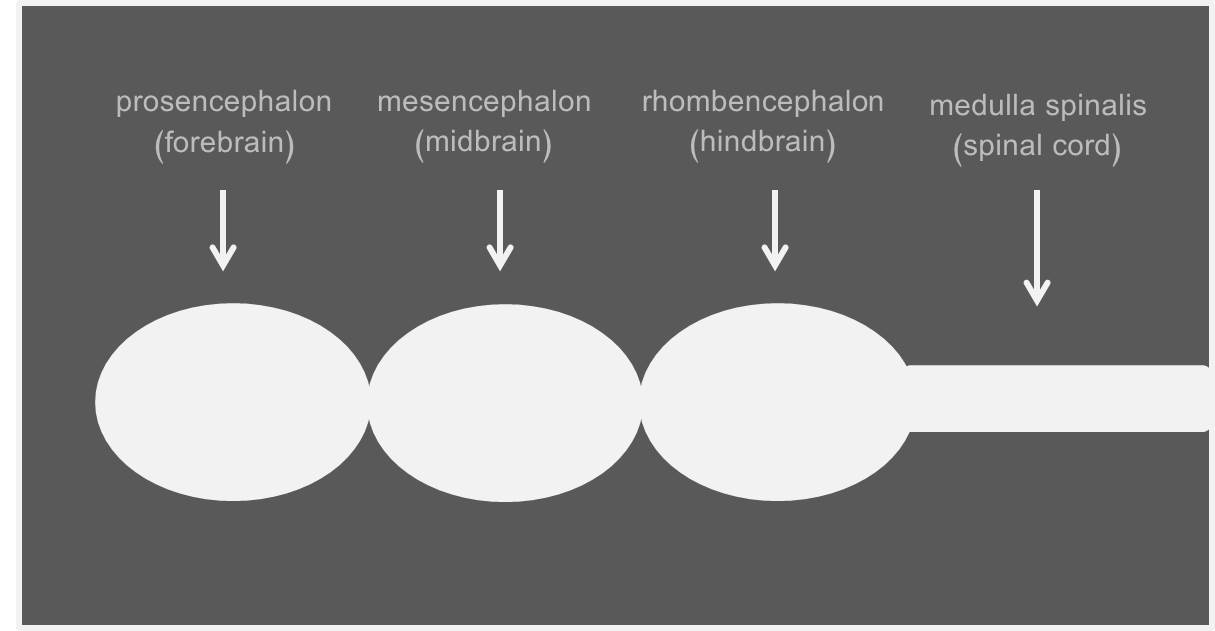
دماغ الفقاريات في بداية ظهوره كان يتكون من ثلاثة حويصلات هي من الخلف للأمام:
الدماغ الخلفي (hindbrain) rhombencephalon
الدماغ الأوسط (midbrain) mesencephalon
الدماغ الأمامي (forebrain) prosencephalon
هذه الحويصلات تظهر عند البشر في المرحلة الجنينية. الرسم التالي يعبر عن الدماغ الجنيني في بداية تكونه:
عند البشر البالغين ما يسمى بالدماغ “الخلفي” هو في الحقيقة “سفلي”، وما يسمى بالدماغ “الأمامي” هو في الحقيقة “علوي”. البشر يمشون على رجلين وليس أربع أرجل، ولهذا السبب الاتجاهات التشريحية لديهم تختلف عن الاتجاهات التشريحية لدي بقية الفقاريات أو ثنائيات الجانب. التسميات المتعلقة بالدماغ تعكس الواقع الحيواني وليس الواقع البشري.
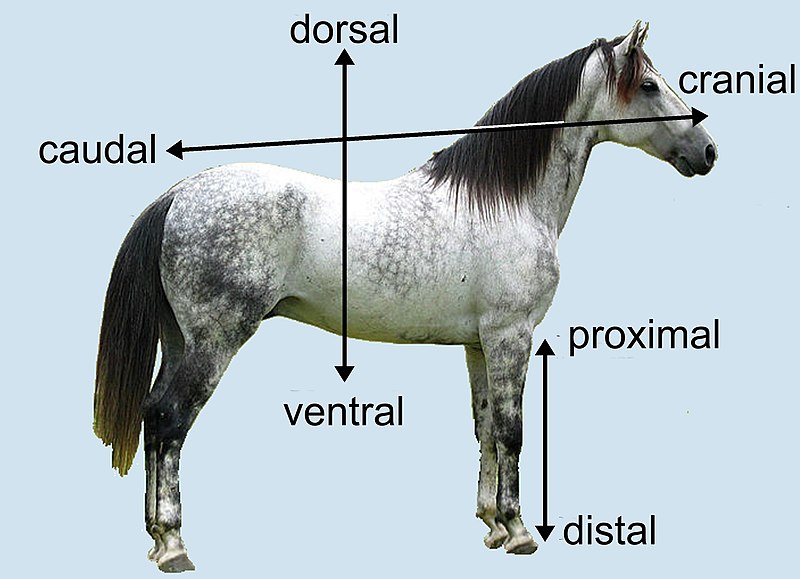
في علم التشريح anatomy الاتجاه الذي يوصل نحو الأنف يسمى الاتجاه “المنقاري” rostral، والاتجاه الذي يوصل نحو الذيل يسمى الاتجاه “الذيلي” caudal. عند الحيوانات التي تمشي على أربعة أرجل الاتجاه المنقاري هو نفسه الاتجاه “الأمامي” anterior، والاتجاه الذيلي هو نفسه الاتجاه “الخلفي” posterior، ولكن عند البشر الاتجاه المنقاري يسمى “العلوي” superior، والاتجاه الذيلي يسمى “السفلي” inferior. الاتجاه الذي يوصل نحو السطح البطني للجسم يسمى الاتجاه “البطني” ventral، والاتجاه الذي يوصل نحو السطح الظهري للجسم يسمى الاتجاه “الظهري” dorsal. عند البشر الاتجاه البطني يسمى “الأمامي” anterior، والاتجاه “الظهري” يسمى “الخلفي” posterior (هذا الكلام لا ينطبق على الأعضاء البشرية ذات المحور الأفقي كالقدم مثلا)
عند الفقاريات الدماغ الخلفي (hindbrain) rhombencephalon يتحول إلى جزأين:
الدماغ النخاعيmyelencephalon (يتحول لاحقا إلى النخاع المستطيل (medulla oblongata
الدماغ التالي metencephalon (يتحول إلى الجسر pons والمخيخ cerebellum)
الدماغ الأوسط (midbrain) mesencephalon عند الثدييات يظل عموما على حاله ولا ينقسم إلى أجزاء.
الدماغ الأمامي (forebrain) prosencephalon يتحول إلى جزأين:
الدماغ البيني diencephalon
الدماغ النهائي telencephalon
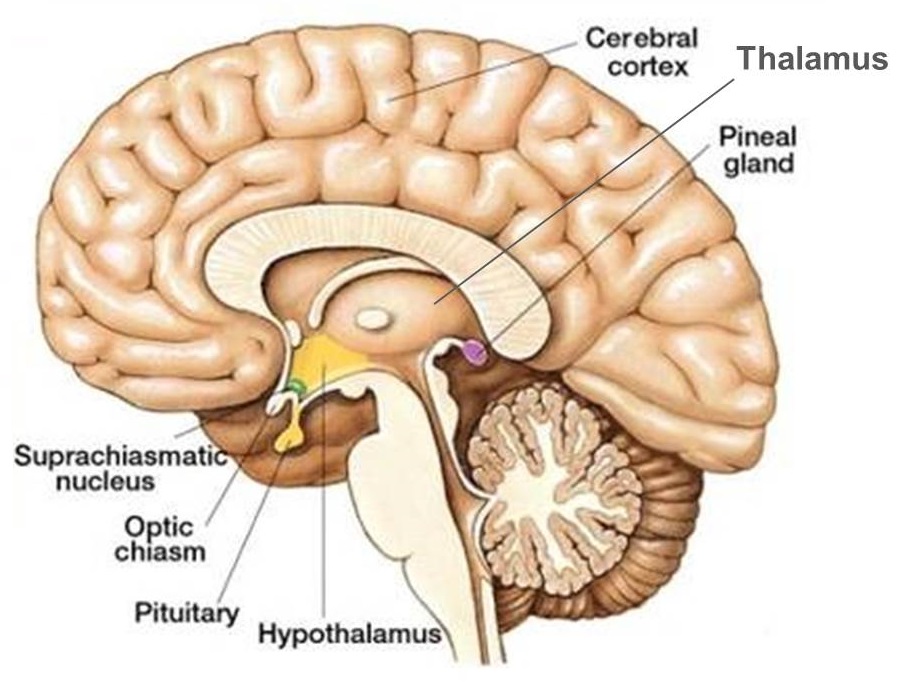
الدماغ البيني diencephalon يتحول إلى جزأين مهمين، الجزء الأول يسمى thalamus (معناه “الحجرة الداخلية” inner chamber). سوف أعرّب كلمة thalamus في هذا المقال إلى “الثَّلَم” (بعض الناس يكتبون “ثلاموس” ولكن من غير الضروري أن نكتب علامة الإعراب اللاتينية –us في الكلمة العربية).
العضو الثاني المهم الذي ينتج من الدماغ البيني هو hypothalamus (سوف أعربه إلى “ما تحت الثلم”).
الدماغ النهائي telencephalon يتحول إلى المخ cerebrum. المخ يتكون من قشرة cortex ومن عقد ganglia أو أنوية nuclei توجد تحت القشرة في بطن المخ.
ما سبب نشوء “الأدمغة”؟
السؤال الذي يطرح نفسه هو لماذا تقسّم دماغ الفقاريات إلى ثلاثة أدمغة (أمامي وأوسط وخلفي)؟
أنا بصراحة لم أقرأ ما يكفي في هذا المجال، وبالتالي ما سأقوله الآن هو مجرد رأي مبني على بعض القراءة المحدودة.
هناك خاصية تميز الحيوانات ثنائية الجانب bilaterian وهي أن أجسامها مقسمة تشريحيا إلى “قطع متتالية”metameres (تسمى أيضا “قطع جسمية” somites).
القطع المتتالية تظهر بوضوح في أجسام الديدان الحلقية:
لاحظ كيف أن جسم الدودة مقسم إلى حلقات متتالية. الديدان هي الحيوانات الأكثر بدائية ضمن مجموعة ثنائيات الجانب، وهذا التقسم إلى قطع متتالية يعبر ربما عن الحالة الأصلية لدى ثنائيات الجانب
القطع المتتالية تظهر أيضا في مفصليات الأرجل arthropods:
لاحظ التقطع المتتالي metamerism لدى أم أربعة وأربعينالتقطع المتتالي لدى العقربالتقطع المتتالي في جسم النحلة
القطع المتتالية المكونة لأجسام مفصليات الأرجل تلتحم مع بعضها لتشكل أجزاء أكبر هي الرأس head والصدر thorax والبطن abdomen.
الأجزاء الكبيرة التي تنتج من التحام القطع المتتالية تسمى tagmata (مفرد tagmata هو tagma ومعناه “المُرَتَّب” something arranged).
في أجسام الحبليات chordates (ومنها الإنسان) هناك “مرتّبات” tagmata شبيهة بالمرتبات الموجودة في أجسام مفصليات الأرجل.
القطع المتتالية metameres أو القطع الجسمية somites تظهر بشكل واضح لدى جنين الإنسان خلال المراحل الأولى من تكوينه:
صورة ظهرية لجنين الإنسانصورة جانبية لجنين الإنسان
جسم جنين الإنسان في بداية تكوينه يكون مقطعا على نحو شبيه بجسم الدودة، ولكن القطع المتتالية تلتحم تدريجيا وتكون “مرتبات” tagmata.
ما هو سبب التحام القطع المتتالية؟ أظن أن السبب هو مبدأ “الاتحاد قوة”. التحام القطع المتتالية مع بعضها يعطيها القدرة على تكوين أعضاء كبيرة ومعقدة. هذا المبدأ هو نفس المبدأ الذي دفع الكائنات وحيدة الخلية لكي تلتحم مع بعضها وتكون الكائنات متعددة الخلايا.
بقايا التقطع المتتالي metamerism تظهر واضحة لدى الإنسان البالغ في العمود الفقري (المكون من عدد كبير من القطع المتتالية التي تسمى الفقرات vertebra). أيضا التقطع المتتالي يظهر واضحا في جدار الصدر:
المناطق التي تقع بين أضلاع القفص الصدري تسمى “المسافات بين الضلعية” intercostal spaces. هذه المسافات لها تكوين متماثل من حيث العضلات والأعصاب والتروية الدموية
التقطع المتتالي في الجهاز العصبي
ظاهرة التقطع المتتالي تظهر واضحة في الجهاز العصبي لثنائيات الجانب. الحبلان العصبيان عند الديدان يحويان عقدا عصبية قطعية segmental ganglia (العقدة العصبية ganglion هي عبارة عن تكتل من الخلايا العصبية). كل قطعة متتالية metamere تحوي عقدتين عصبيتين على اليمين وعلى الشمال، ومن كل عقدة عصبية تخرج أعصاب محركة نحو عضلات القطعة المتتالية التي تحوي العقدة. الأعصاب تقبّض عضلات القطع المتتالية بشكل متناسق وهذا ما يحرك الدودة.
في رأس الدودة توجد عقدتان كبيرتان تسميان “العقدتان المخيتان” cerebral ganglia. هاتان العقدتان هما “دماغ الدودة”. هاتان العقدتان تتصلان مع عدد من أعضاء الحس البدائية (هناك لدى دودة الأرض “عين” بدائية تستطيع أن تفعل شيئا واحدا هو تحديد وجود الضوء من عدمه، وهناك حاسة شم وذوق بدائية تستطيع التعرف على وجود بعض المواد الكيميائية، وهناك أعضاء حسية في الجلد تستطيع الشعور بالحرارة والاهتزاز وإرسال إشارات إلى العقد المخية لإبلاغها بذلك). العقدتان المخيتان تستفيدان من معطيات أعضاء الحس وترسلان إشارات إلى العضلات لتحريك الدودة بعيدا عن الضوء والاهتزاز والأمور الأخرى التي قد تهدد حياة الدودة.
هذا الجهاز العصبي البدائي الموجود لدى الديدان هو على ما أظن أساس الأجهزة العصبية المعقدة الموجودة لدى حيوانات أخرى أكثر تطورا.
مفصليات الأرجل تملك جهازا عصبيا شبيه بالجهاز العصبي الدودي، ولكن في مفصليات الأرجل هناك اندماج للعقد العصبية في وسط البطن:
الجهاز العصبي لنحلة بالغة (B) وليرقة النحل (A)
ظاهرة الاندماج بين الحبلين العصبيين هي موجودة أيضا في الحبليات، ولكن الحبل العصبي لدى الحبليات هو ظهري وليس بطنيا كما في مفصليات الأرجل.
دماغ مفصليات الأرجل هو عبارة عن عقدة عصبية كبيرة نشأت من اندماج أزواج العقد العصبية الثلاثة الأولى (المسؤولة عن تعصيب القطع المتتالية metameres الثلاث الأولى في جهة الرأس).
دماغ مفصليات الأرجل ينقسم إلى ثلاثة أجزاء، الجزء الأمامي يسمى “المخ الأول”protocerebrum وهو يتصل مع أعضاء الرؤية (الحشرات عموما لديها خمسة عيون: عينان مركبتان compound eyes تضمان عددا كبيرا من العدسات سداسية الأضلاع، وثلاث عيون بسيطة simple eyes تضم كل منها عدسة واحدة).
العينان المركبتان على الجانبين، والعيون البسيطة في الوسط
الجزء الثاني من دماغ مفصليات الأرجل هو “المخ الثاني” deutocerebrum الذي يتصل مع قرون الاستشعار antennae (المسؤولة عن حاسة الشم). الجزء الثالث من دماغ مفصليات الأرجل هو “المخ الثالث” tritocerebrum الذي لا أعلم ما هي وظيفته (من الممكن حسب هذا الرابط أنه مسؤول عن استقبال وإرسال البيانات إلى أعضاء الجسم الحشوية visceral).
إلى الأسفل من العقدة المخية الكبيرة هناك عقدة كبيرة مكونة من اندماج ثلاثة أزواج من العقد العصبية تسمى “العقدة تحت المريئية” subesophageal ganglion. هذه العقدة مسؤولة عن تعصيب الفكين والفم بما في ذلك الغدد اللعابية وربما حس الذوق.
أحاسيس مفصليات الأرجل هي أكثر تعقيدا من أحاسيس الديدان. وظيفة العيون البسيطة (التي تسمى “العيون الصغيرة” ocelli) هي تحديد وجود الضوء وشدته. وظيفة العيون المركبة هي رصد حركة الأجسام في محيط الحشرة (وليس “رؤية” الأجسام بالمعنى البشري للكلمة، لأن العيون المركبة هي ليست مناسبة لتكوين صورة للأجسام). قرون الاستشعار هي قادرة على الإحساس بجزيئات المواد الكيميائية المحمولة في الهواء (أي أنها قادرة على الشم). في الفم توجد على ما يبدو أعضاء حس قادرة على الإحساس بجزيئات المواد الكيميائية الذائبة (أي التذوق). جلد الحشرات يحوي مستشعرات للاهتزاز، ويبدو أن هناك في الصدر أو البطن مستشعرات قادرة على الإحساس باهتزاز جزيئات الهواء (أي أنها قادرة على السمع). هناك أيضا في أجسام الحشرات مستشعرات قادرة على الإحساس بالضغط والشد، أي أن الحشرات تملك ما يسمى بـ”الحس الذاتي” proprioception (الإحساس بالحركة في أعضاء الجسم).
التقطع المتتالي في الجهاز العصبي للفقاريات
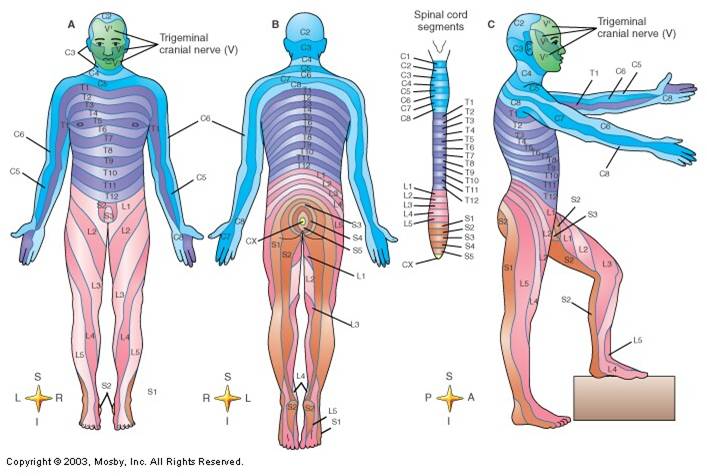
ظاهرة التقطع المتتالي في الجهاز العصبي تظهر بشكل واضح لدى الفقاريات في العضو المسمى بالحبل الشوكي (الحبل الشوكي هو نفسه الحبل العصبي ولكن تسميته بـ”الشوكي” لدى الفقاريات تعود إلى كونه محصورا بين النتوءات العظمية “الشوكية” التي تخرج من فقرات العمود الفقري). الحبل الشوكي لدى الفقاريات يبدو في الشكل ملتحما وخاليا من العقد القطعية segmental ganglia، ولكن لو نظرنا إلى الطريقة التي يعمل بها هذا الحبل فسيتبين لنا أنه مقطّع بامتياز.
الحبل الشوكي هو مقسم وظيفيا إلى قطع تسمى spinal cord segments. كل قطعة من هذه القطع تنتج عصبين شوكيين spinal nerves (واحد من جهة اليمين وآخر من جهة الشمال). العصبان الشوكيان يخرجان من المسافات بين نواتئ الفقرات.
منظر من الجهة اليسرى للعمود الفقري يبين خروج أعصاب شوكية من بين نواتئ فقرات العمود الفقري
الأعصب الشوكية هي نفسها مقطعة وظيفيا. كل عصب شوكي يحوي قسما حسيا sensory وقسما محركا motor. الألياف العصبية الحسية المسؤولة عن نقل الإحساس من الجلد هي مقطعة وظيفيا على نحو واضح جدا.
المناطق المخططة على الجلد تسمى “قطاعات جلدية” dermatomes. هناك عصب شوكي واحد مسؤول عن نقل الإحساس من كل قطاع جلدي على يمين الجسم وشماله. على الصورة تظهر قطع الحبل الشوكي مع أسمائها. القطع التي تبدأ أسماؤها بالحرف C هي القطع الرقبية cervical، والقطع التي تبدأ أسماؤها بالحرف T هي القطع الصدرية thoracic، والقطع التي تبدأ أسماؤها بالحرف L هي القطع القَطَنية lumbar، والقطع التي تبدأ أسماؤها بالحرف S هي القطع العَجُزِية sacral، والقطعة المسماة CX هي القطعة العصعصية coccygeal. لاحظ أن القطاعات الجلدية الأخيرة تقع حول الشرج في المنطقة التي كان يخرج منها الذيل سابقا. بالنسبة للقطاعات الجلدية الوجهية (التي تحمل الرمز V) فهي متصلة مع أعصاب تخرج من جذع الدماغ.
لو رسمنا القطاعات الجلدية dermatomes على جسم الإنسان بواسطة قلم فإن شكل جسمه سيصبح شبيها بجسم الدودة الحلقية.
الأعصاب الشوكية الحسية حافظت في وظيفتها إلى حد كبير على التقطع المتتالي الموروث من الأزمنة القديمة (من زمن ثنائيات الجانب الأولى urbilaterians). سبب ذلك هو أن جلد الإنسان لم يتكتل ويشكل أعضاء كبيرة. المبدأ الذي يعمل عليه الجلد البشري لا يختلف كثيرا عن المبدأ الذي يعمل عليه جلد الديدان.
بالنسبة للأعصاب الشوكية المحركة (المسؤولة عن تحريك العضلات) فهي لم تحافظ في وظيفتها لدى البشر على التقطع المتتالي المشاهد لدى الديدان. السبب هو أن النسيج العضلي في جسم الإنسان متكتل على هيئة عضلات كبيرة عابرة لحدود القطع المتتالية أو القطع الجسمية. الأعصاب المحركة اضطرت للتكتل لكي تجاري العضلات. نفس الأمر ينطبق على الأعصاب الحشوية visceral المسؤولة عن تعصيب الأعضاء التي تقع في جوف الجسم (هذه الأعصاب هي فروع من الأعصاب الشوكية). كثير من الأعضاء الحشوية (القلب، الرئتان، إلخ) هي عبارة عن كتل نسيجية كبيرة عابرة لحدود القطع المتتالية، ولهذا السبب أعصابها هي عبارة عن كتل من الألياف العصبية التي تعود أصولها لعدة عقد قطعية (ولكن لو نظرنا إلى الأمعاء فسنجد أن أعصابها حافظت على تقطعها المتتالي إلى حد ما، لأن الأمعاء هي أقل تكتلا من غيرها من الأعضاء الحشوية).
الأمعاء حافظت على تقطعها المتتالي سواء من حيث التروية الدموية أو التصريف اللمفي أو التعصيب أو حتى التكوين العضلي. هذا ليس مفاجئا لأن الطريقة التي تتحرك بها الأمعاء هي فعليا نفس الطريقة التي تتحرك بها الديدان. الأمعاء هي جزء من الأحشاء الفقارية حافظ إلى حد كبير على طابعه الأصلي الشبيه بالديدان
التقطع المتتالي في الدماغ الفقاري
من كل المقدمة السابقة أردت أن أصل إلى السبب الذي أدى لتكوين الأدمغة الفقارية الثلاثة (الدماغ الخلفي والأوسط والأمامي). دماغ الفقاريات هو في الأصل مجرد امتداد للحبل الشوكي، ولا بد أن دماغ الفقاريات كان في الأصل مكونا من قطع أو عقد عصبية متتالية على غرار الحبل الشوكي. لاحقا حصلت عملية التحام بين عقد دماغ الفقاريات على غرار ما هو مشاهد لدى مفصليات الأرجل. لو قارنا دماغ الفقاريات مع دماغ مفصليات الأرجل فيمكننا أن نتخيل السبب الذي أدى لالتحام العقد الدماغية المتتالية مع بعضها: هدف الالتحام كان تكوين عقد دماغية كبيرة متخصصة في التعامل مع أعضاء الحس، خاصة تلك الموجودة في الرأس (خاصة أعضاء الرؤية والشم).
نشوء الدماغ cephalization ترافق مع تجمع عدد من أعضاء الحس المهمة في منطقة الرأس. هذا الأمر لم يكن مصادفة. من الواضح أن هناك ارتباطا بين تجمع أعضاء الحس في منطقة الرأس وبين نشوء الدماغ.
القسم الأمامي من دماغ مفصليات الأرجل هو متخصص في التعامل مع العيون. القسم الثاني من دماغ مفصليات الأرجل هو متخصص في التعامل مع قرون الاستشعار المسؤولة عن حاسة الشم. القسم الثالث من دماغ مفصليات الأرجل هو مجهول الوظيفة ولكن من الممكن أن وظيفته هي ربط حسي الرؤية والشم مع الأحاسيس الأخرى القادمة من مناطق الجسم السفلى.
لو نظرنا إلى أدمغة الفقاريات البدائية فسنجد توزيعا مماثلا للوظائف:
هذا الرسم يعبر عن أدمغة الفقاريات البدائية التي تجمع عادة تحت عنوان “الأسماك”. القسم الأمامي من أدمغة الأسماك هو المخ cerebrum. شكل المخ هو معقد بعض الشيء. المخ ينشأ خلال المرحلة الجينية من القسم الأمامي من الحبل العصبي (الحبل العصبي عند الأجنة يسمى “الأنبوب العصبي” neural tube). الشكل A في الصورة التالية هو لمقطع عرضي في القسم الأمامي من الأنبوب العصبي خلال المرحلة الجنينية:
الشكل A هو مقطع في الأنبوب العصبي حيث سيتكون المخ. سقف الأنبوب العصبي (المظلل على الرسم) سوف ينفتح وهذا سوف يؤدي لنشوء شكل شبيه بالحرف V. عند الأسماك ذراعا الحرف V سوف ينثنيان نحو الخارج كما يظهر في الشكل C، وعند رباعيات الأرجل tetrapods ذراعا الحرف V سوف ينثنيان نحو الداخل كما يظهر في الشكل B.
على الصورة تظهر مناطق مرقمة بأرقام من 1 إلى 5. هذه المناطق لها أسماء. الشكل التالي يوضح أسماءها:
طبعا هذا الشكل يعبر عن مخ رباعيات الأرجل وليس الأسماك. المناطق العلوية (بالأحرى الظهرية dorsal) من المخ تسمى pallium (هذه المناطق تحمل الأرقام 3 و4 و5 على الصورة السابقة). المناطق السفلية (البطنية ventral) من المخ تسمى subpallium (الرقم 2) و septum (الرقم 1).
الأقسام المخية التي تسمى pallium هي مشتقة من النصف الظهري من الأنبوب العصبي.
معنى كلمة pallium هو الرداء الذي يرتديه البابا في هذه الصورة، أو الرداء الذي يرتديه هذا الرجل. أنا سأترجم كلمة pallium إلى “رداء” رغم أن الترجمة الأصح هي ربما “عباءة”.
على الرسومات التي في الأعلى يظهر أن “الرداء” pallium له ثلاثة أقسام:
الرداء الإنسي medial pallium
الرداء الظهري dorsal pallium
الرداء الوحشي lateral pallium
في التشريح كلمة “وسطي” medial (تعريبها “إنسي”) تعني الاتجاه الذي يوصل نحو المحور السهمي sagittal (أي أنها تعني اتجاه اليسار في النصف الأيمن من الجسم واتجاه اليمين في النصف الأيسر من الجسم). كلمة “جانبي” lateral (تعريبها “وحشي”) تعني الاتجاه الذي يبتعد عن المحور السهمي (اتجاه اليمين في النصف الأيمن من الجسم واتجاه اليسار في النصف الأيسر من الجسم). طبعا عندما يقال في التشريح “يمين” أو “يسار” فالمقصود هو الاتجاهات بالنسبة للكائن نفسه (مثلا “اليد اليمنى” لشخص معين هي اليد التي يراها هذا الشخص على يمينه).
الرداء الإنسي medial pallium لدى جميع الفقاريات هو مسؤول ربما عن الذاكرة (تقليديا كان بعض الناس يقولون أن الأسماك ليس لديها ذاكرة، ولكن حاليا هذا الكلام لم يعد مقبولا حسب ما أعلم).
الرداء الوحشي lateral pallium هو مسؤول عن إدراك حاسة الشم لدى جميع الفقاريات (وحاسة الذوق أيضا على ما أظن، ولكن بالنسبة للأسماك حاستا الشم والذوق هما حاسة واحدة).
الرداء الظهري dorsal pallium هو مجهول الوظيفة لدى الأسماك والفقاريات البدائية، ولكن من الممكن أن وظيفته هي الربط بين البيانات الحسية والذاكرة.
الشكل التالي يبين التوزع النهائي لمناطق الرداء في مخ سمكة عظمية (في الأسفل) ومخ حيوان قارض (من الثدييات) (في الأعلى):
القسم المخي الجنيني الذي يقع مباشرة تحت الرداء (المسمى “تحت الرداء” subpallium) يتحول في النهاية إلى مجموعة من الأعضاء المهمة التي تسمى “العقد القاعدية” basal ganglia. العقد القاعدية توجد في قلب المخ، ووظيفتها في رأيي مرتبطة بالتعلم learning (أي إضافة معلومات جديدة إلى الذاكرة).
الدراسات أثبتت قدرة الأسماك على التعلم. الأسماك تستطيع أن تتعلم ما هي الكائنات المفترسة لكي تتجنبها.
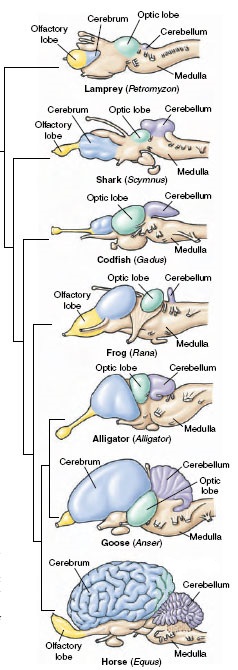
المخ لدى جميع الفقاريات (تحديدا مناطق القشرة المخية الناتجة من الرداء الوحشي lateral pallium) هو متصل مع البصلتين الشميتين olfactory bulbs اللتان تنقلان حس الشم من أعضاء الشم. حسب علمي فإن المناطق المخية المسؤولة عن إدراك حاسة الشم تقع لدى جميع الفقاريات إلى الأمام من المناطق المخية المسؤولة عن الذاكرة والتعلم. لهذا السبب لو نظرنا إلى صور أمخاخ الفقاريات فسنجد أن المناطق المخية المسؤولة عن إدراك حاسة الشم تقع دائما في مقدمة المخ.
المناطق المسؤولة عن إدراك حاسة الشم ملونة باللون الأصفر. الكائنات هي من الأعلى إلى الأسفل: lamprey (سمكة بلا فكين)، سمك القرش (سمكة غضروفية)، codfish (سمكة عظمية)، ضفدع (برمائي)، تمساح (زاحف)، إوزة (طائر)، حصان (ثديي).
من يدقق في الصورة أعلاه سوف يلاحظ أن نسبة اللون الأصفر (حاسة الشم) تقل بشكل تدريجي في أمخاخ الفقاريات كلما ازداد تطور الحيوان الفقري، وأما نسبة اللون الأزرق (المناطق المسؤولة عن الذاكرة والتعلم) فهي تزداد بشكل تدريجي كلما ازداد تطور الحيوان الفقري. عند حيوان الـ lamprey اللون الأزرق يغطي أقل من نصف المخ، أي أن معظم مخ هذا الحيوان هو مخصص لحاسة الشم.
هذا يعيدنا إلى الفكرة التي بدأت بها حديثي حول دماغ الفقاريات. أنا قارنت بين دماغ الفقاريات وبين دماغ مفصليات الأرجل، وقلت أن نشوء “الأدمغة” (الأمامية والوسطى والخلفية) كان مرتبطا بتطور الحواس. أنا أظن أن المخ (الدماغ النهائي telencephalon) نشأ لدى الفقاريات في الأصل بهدف التعامل مع حاسة الشم. وظائف الذاكرة والتعلم هي تطورات لاحقة وليست الهدف الأصلي الذي ظهر لأجله المخ.
حاليا لو نظرنا إلى مخ الإنسان فسنجد أن المناطق المسؤولة عن معالجة حاسة الشم هي صغيرة للغاية ولا تكاد تمثل شيئا من المخ، ولكن المفارقة هي أن المخ نشأ في الأصل بسبب حاسة الشم. التعامل مع حاسة الشم تطلب قدرات عصبية كبيرة نسبيا، وهذا هو ما حفز العقد العصبية القطعية (الموروثة من الحيوانات ثنائية الجانب bilaterian) على التكتل وتشكيل المخ.
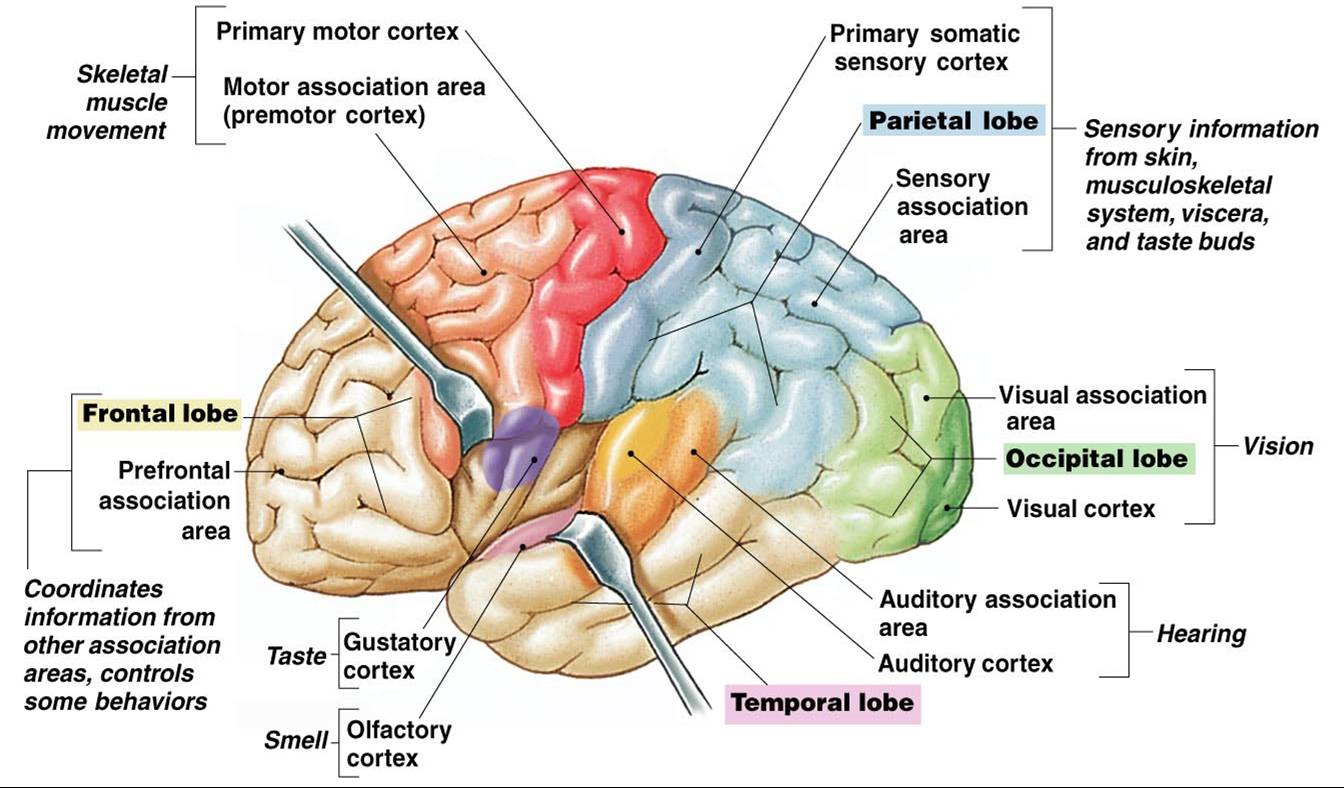
رسم يبين موقع القشرة الشمية olfactory cortex والقشرة الذوقية gustatory cortex في المخ البشري
بنفس الطريقة يمكن في رأيي أن نفسر ظهور بقية أجزاء الدماغ لدى الفقاريات. الجزء التالي للدماغ النهائي (المخ) في دماغ الفقاريات هو الدماغ البيني diencephalon. ما هي يا ترى الوظيفة الأصلية للدماغ البيني؟
الدماغ البيني لدى الإنسان والثدييات هو من أعقد أجزاء الدماغ (وأهمها)، ولكن الوظيفة الأصلية لهذا الدماغ كانت بسيطة: الدماغ البيني كان في الأصل مسؤولا عن التعامل مع حس البصر.
أهم جزء في العينين هو الشبكية retina التي تستقبل الضوء وتحوله إلى إشارات عصبية. الشبكية تنشأ من عضوين جنينيين هما “الحويصلان البصريان” optic vesicles اللذان ينبتان من الدماغ البيني.
حس البصر ينتقل من شبكيتي العينين عبر العصبيين البصريين optic nerves ويمر عبر التصالب البصري optic chiasm ويصل إلى “الجسمين الركبيين الوحشيين” lateral geniculate bodies اللذين يقعان على الجهة الظهرية من الثلم thalamus. الجسمان الركبيان الوحشيان يرسلان بيانات حس البصر في اتجاهين:
الاتجاه الأهم هو نحو القشرة البصرية visual cortex في مؤخرة المخ
الاتجاه الأقل أهمية هو نحو “التليلتين العلويتين” superior colliculi الموجودتين على الجهة الظهرية للدماغ الأوسط (midbrain) mesencephalon
القشرة البصرية في المخ البشري هي اختراع حديث ظهر لأول مرة لدى الثدييات. الفقاريات السابقة للثدييات لم تكن تملك هذه القشرة. في تلك الكائنات بيانات حس البصر كانت تنتقل من المنطقة الظهرية للثلم thalamus نحو المنطقة الظهرية من الدماغ الأوسط التي تسمى “السقف” tectum.
سقف الدماغ الأوسط كان ربما أهم جزء في دماغ الفقاريات الباكرة. عند الأسماك سقف الدماغ الأوسط يشكل نصفي كرة على غرار نصفي الكرة المخيين، والمفارقة هي أن نصفي الكرة السقفيين هما أكبر من نصفي الكرة المخيين:
دماغ سمكة عظمية. لاحظ أن سقف الدماغ الأوسط tectum هو أكبر أجزاء الدماغمنظر ظهري لدماغ سمكة عظمية
نصفا الكرة السقفيان المسؤولان عن معالجة حس البصر يسميان “الفصان البصريان” optic lobes. هذا الفصان ما زالا موجودين لدى الثدييات ولكنهما تقزما وأصبحا مجرد حدبتين صغيرتين لا يمكن رؤيتهما إلا بصعوبة.
منظر إنسي (من الداخل) للنصف الأيسر من مخ بشري. السهم يشير إلى بقايا الفص البصري في سقف الدماغ الأوسط. في الدماغ البشري هذا الفص يسمى “التليلة العلوية” superior colliculus (كلمة colliculus في اللاتينية هي تصغير لـ collis التي تعني “تلة”).
عند البشر هناك ثلاثة أماكن يتم فيها معالجة البيانات البصرية القادمة من شبكيتي العينين:
ظهر الثلم (الجسمان الركبيان الوحشيان lateral geniculate bodies)
ظهر (سقف) الدماغ الأوسط (التليلتان العلويتان superior colliculi)
ظهر المخ (القشرة البصرية visual cortex)
كل منطقة من هذه المناطق الثلاث تستخدم البيانات البصرية لتحقيق أهداف مختلفة. ظهر الثلم يستفيد من البيانات البصرية لتحديد مواقع الأجسام المحيطة بالإنسان ورصد حركتها في حال تحركت. سقف الدماغ الأوسط يستفيد من البيانات البصرية لتحريك عضلات الجسم في حال كانت هناك ضرورة لذلك. عند البشر السقف هو مسؤول عن عدد قليل من الحركات العضلية (هي تحديدا بعض الحركات اللاإرادية أو الأوتوماتيكية، خاصة حركات العينين)، ولكن في الفقاريات القديمة السقف كان يلعب دورا جوهريا في تحريك الجسم عموما.
القشرة البصرية عند البشر تستفيد من البيانات البصرية بطرق عديدة تدخل ضمن ما يسمى بالقدرة الاستعرافية cognition. القشرة البصرية هي المسؤولة عما يسمى بالإدراك البصري visual perception (الذي يسميه الناس “الرؤية” vision). لو تعرضت القشرة البصرية للتلف فإن الإنسان سيفقد الرؤية، أي أنه سيصبح أعمى (هذا يحدث بسبب الجلطات الدماغية أو الحوادث التي تقطع التروية الدموية عن القشرة البصرية).
ولكن حتى بعد تلف القشرة البصرية وإصابة الإنسان بالعمى فإن حاسة البصر تظل تعمل. هذا الأمر قد يبدو غريبا لكثير من الناس، لأن الناس يعتقدون أن الرؤية أو “الإدراك البصري” هو نفسه حاسة البصر.
هناك ظاهرة تسمى “الرؤية العمياء” blindsight. هذه الظاهرة تشاهد لدى الأشخاص المصابين بالعمى نتيجة لتلف القشرة البصرية. هؤلاء الناس لا يرون شيئا (لأنهم عميان)، ولكنهم رغم ذلك “يشعرون” بالأشياء الموجودة حولهم و”يشعرون” بحركة هذه الأشياء في حال تحركت.
تفسير هذه الظاهرة هو أن ظهر الثلم يقوم بمعالجة البيانات البصرية ويستفيد منها لتحديد مواقع الأشياء وحركتها. بما أن القشرة البصرية تالفة فالإنسان لا “يرى” الأشياء التي يدركها الثلم.
مفهوم “الرؤية العمياء” هو نفس مفهوم الرؤية لدى الحشرات. العيون المركبة compound eyes لدى الحشرات هي عمليا غير صالحة للرؤية، ولكن هذه العيون تفيد في تحديد مواقع الأشياء وحركتها. الحشرات تستطيع أن تحدد مواقع الأجسام من حولها وتستطيع أن تحدد بدقة ما إذا كانت هذه الأجسام متحركة أم غير متحركة، رغم أن الحشرات لا “ترى” أي شيء.
مفهوم حاسة البصر هو أوسع من مفهوم “الرؤية”. حاسة البصر هي موجودة لدى الحيوانات منذ الزمن الذي ظهرت فيه ثنائيات الجانب لأول مرة قبل مئات ملايين السنين، ولكن “الرؤية” هي شيء حديث ظهر لأول مرة لدى الثدييات.
الديدان تملك عيونا بدائية تسمح لها بتحديد وجود الضوء من عدمه، والحشرات تملك أيضا هذا النوع من العيون. الحشرات تملك نوعين من العيون، عينان مركبتان وثلاث عيون بسيطة.
العيون البسيطة محاطة بدائرةالعيون البسيطة ظاهرة خلف قرني الاستشعار
أنا أظن أن الثلم thalamus (والدماغ البيني عموما) نشأ لأول مرة بهدف التعامل مع عيون بدائية شبيهة بالعيون المركبة لدى الحشرات. هذه العيون لم تكن قادرة على الرؤية ولكنها كانت قادرة على تحديد مواقع الأجسام وحركتها.
في الحقيقة هناك دلائل على أن الدماغ البيني القديم كان يتعامل أيضا مع عيون بسيطة شبيهة بالعيون البسيطة الموجودة لدى الحشرات (التي تدرك شدة الضوء). من يريد أن يتوسع في هذا الموضوع يمكنه أن يرجع إلى هذا الرابط.
العين البسيطة (تسمى العين الجدارية parietal eye) هي موجودة لدى الكثير من الفقاريات (الأسماك، البرمائيات، الزواحف)، وهي تؤدي نفس وظيفة العيون البسيطة لدى الحشرات.
العين الجدارية parietal eye هي الدائرة الفضية الصغيرة بين عيني الضفدعالعين الجدارية بين عيني سحلية
“العين الجدارية” أو “العين الثالثة” تتصل بشكل مباشر مع عضو يسمى “ما فوق الثلم” epithalamus. هذا العضو يقع على الجهة الظهرية للثلم وهو جزء من الدماغ البيني. “ما فوق الثلم” يتصل مع “الغدة الصنوبرية” pineal gland التي تقع في مؤخرة الثلم.
الغدة الصنوبرية تلعب دورا في تنظيم مواعيد النوم والاستيقاظ، أو ما يسمى “الإيقاع اليومي” circadian rhythm. قديما كانت الغدة الصنوبرية تعتمد على العين الجدارية لتقدير شدة ضوء الشمس. عندما يكون الضوء الساقط على العين الجدارية شديدا فهذا يعني أن الوقت هو نهار، وبالتالي الحيوان يجب ألا ينام (إذا كانت حياته نهارية)، وعندما تنخفض شدة الضوء الساقط على العين الجدارية فهذا يعني أن الوقت هو ليل، وبالتالي الحيوان يجب أن ينام. الغدة الصنوبرية تلعب أيضا دورا في تنظيم درجة حرارة الجسم.
منظر إنسي (من الداخل) لنصف الكرة المخية الأيمن
العين الجدارية اختفت من عند الثدييات، والغدة الصنوبرية عند الثدييات هي حسب علمي عضو قليل الأهمية أو شبه أثري vestigial. تنظيم الإيقاع اليومي ودرجة الحرارة عند الثدييات يتم بالدرجة الأولى في منطقة “ما تحت الثلم” hypothalamus، التي هي منطقة مهمة تقع على الجانب البطني الأمامي من الثلم (وتتصل مع الجهاز الغدي الداخلي endocrine system عبر تحكمها بالغدة النخامية pituitary gland). “ما تحت الثلم” يعتمد على إشارات بصرية واردة مع العصبين البصريين لتقدير شدة ضوء الشمس. عينا الثدييات الجانبيتان تطورتا وأصبحتا قادرتين على تحديد شدة ضوء الشمس، وبالتالي لم تعد هناك حاجة للعين الجدارية (التي تسمى أيضا “العين الصنوبرية” pineal eye).
الخلاصة هي أن الدماغ البيني diencephalon كان في الأصل يشرف على نوعين من العيون: العين الجدارية والعينان الجانبيتان. هذه في رأيي (وفي رأي غيري) هي الوظيفة الأصلية للدماغ البيني التي أدت لنشوئه من الأساس.
بالنسبة للدماغ الأوسط midbrain) mesencephalon) فهو يؤدي عددا كبيرا من الوظائف في الدماغ البشري، وما لفتني في وظائفه هو أنها معقدة وذات أهمية محورية لعمل الدماغ البشري عموما. أنا لا أظن أن الوظيفة الأصلية للدماغ الأوسط كانت مجرد الإشراف على حاسة معينة. في رأيي أن الدماغ الأوسط كان منذ البداية مسؤولا عن ربط الإشارات الحسية مع الإشارات التحريكية بهدف تحفيز السلوك لدى الحيوان، أي أنه كان يلعب دورا شبيها بدور “اللوحة الأم” motherboard في أجهزة الكومبيوتر. ولكن هذا التشبيه قد تكون فيه مبالغة، لأننا لو نظرنا إلى مفصليات الأرجل مثلا فسنجد أن جهازها العصبي يخلو من “لوحة أم”. الجهاز العصبي لدى مفصليات الأرجل والديدان هو إلى حد كبير غير مركزي. الحشرة يمكنها أن تعيش لفترة طويلة حتى بعد إزالة دماغها (عقدها المخية cerebral ganglia).
الجهاز العصبي لدى الحيوانات هو من الأصل غير مركزي. تركيب أجسام الحيوانات هو قائم من الأساس على اللامركزية. السبب بسيط وأنا شرحته في الأعلى بشكل مفصل: أجسام الحيوانات تكونت وفق مبدأ “التكتل ثم التخصص”. الحياة في الأصل كانت تقوم على مبدأ الخلية الواحدة. بعد ذلك ظهرت الكائنات متعددة الخلايا بسبب تكتل الخلايا الأحادية مع بعضها. في أجسام الكائنات متعددة الخلايا حدثت عملية تخصص: كل مجموعة من الخلايا صارت تؤدي وظيفة محددة، وهذا ما أدى إلى نشوء الأنسجة الخلوية المتخصصة (نسيج عضلي ونسيج عصبي ونسيج جلدي إلخ). بعد ذلك ظهرت الحيوانات ذات التقطع المتتالي metamersim. المبدأ الذي قامت أجسام هذه الحيوانات على أساسه هو نفس مبدأ التكتل آنف الذكر—أجسام هذه الحيوانات هي عبارة عن تكتل لقطع متتالية metameres ذات تركيب نسيجي متماثل. الخطوة التالية هي التخصص مجددا—كل مجموعة من القطع المتتالية كونت “مرتبا” tagma يؤدي وظائف خاصة.
أجسام الحيوانات نشأت وفق طريقة لا علاقة لها بالمركزية. لا توجد جهة مركزية معينة أشرفت على تطوير أجسام الحيوانات وقررت أن توزع الأدوار على أجزاء الجسم المختلفة.
فكرة “الإدارة المركزية” هي في رأيي فكرة لا علاقة لها بعالم الطبيعة. هذه الفكرة هي من بنات أفكار البشر. هذه الفكرة ظهرت خصوصا بعد ظهور المجتمعات الإقطاعية، ونشوؤها في رأيي كان مرتبطا بالجهل وليس فقط بنوازع التسلط لدى البشر.
البشر بجهلهم لطالما اعتقدوا أنهم يستطيعون أن يسيروا الطبيعة وفق هواهم. كل الأفكار المركزية تقوم على منطق التحكم بالطبيعة أو البيئة. البشر كثيرا ما يكرهون العالم الذي يعيشون فيه ويعتقدون أنه فوضوي أو غير عادل إلخ، ولهذا السبب هم كثيروا ما يقررون أن يفرضوا سيطرتهم على الطبيعة وأن يسيروها وفق هواهم.
المراقبة العلمية للطبيعة التي نعيش فيها تبين أنها معقدة للغاية وتفوق بكثير قدرتنا على الإدراك والفهم. نحن بالكاد نفهم ما يجري من حولنا (سواء من الناحية البيولوجية أو المناخية أو الاجتماعية أو الاقتصادية إلخ). معظم العلوم التي تدرس الطبيعة والبيئة والمجتمع هي علوم قاصرة ولا تقدم فهما كاملا وأكيدا لما يجري من حولنا. نحن لسنا قادرين على فهم التفاعلات والتغيرات المعقدة التي تجري في الطبيعة من حولنا، فما بالك بأن نحاول التحكم بالطبيعة وتسييرها وفق عاطفتنا.
البشر بجهلهم لم يستوعبوا ذلك. هم في الغالب انطلقوا من أساس عاطفي وظنوا أن مَلِكهم يستطيع أن يتحكم بالطبيعة كما يتحكم بأفراد المجتمع (لأن الملك هو شخص ذو سلطة مطلقة، أو هو إله كما كان القدماء يعتقدون).
ظهور الأفكار المركزية في الإدارة هو مرتبط تاريخيا بظهور التقديس والتأليه للملوك. الحضارات الأولى التي قامت في الشرق الأوسط كانت تربط بين عمل الحكومات المركزية وعمل الآلهة. بالنسبة لهم قرارات الحكومات كانت قرارات إلهية مدعومة من الآلهة. ليس من الغريب إذن أن الفكر المركزي في الإدارة تعاظم لديهم إلى درجة أنه في النهاية أدى لانهيار حضاراتهم (الحضارة السومرية انهارت في حدود عام 2000 قبل الميلاد بسبب النظام الإداري والاقتصادي الفاشل الذي كان قائما على المركزية المفرطة).
الدول الاشتراكية التي رأيناها في القرن العشرين هي مثال آخر على فشل النظام المركزي. الاتحاد السوفييتي كان دولة هائلة الحجم والقدرات، ولكنه انهار في نهاية الثمانينات على نحو مفاجئ ومضحك.
النظام السوري بأفكاره المركزية والاشتراكية جعل سورية إحدى أفشل وأسوأ الدول في العالم.
الإمبراطوريات العظيمة التي استمرت لقرون عديدة كانت تقوم على اللامركزية. الإيرانيون والرومان والعرب كانوا يطبقون اللامركزية في الإدارة والثقافة، وهذا هو سبب ازدهار حضاراتهم وتأثيرها الواسع. أيضا في عصرنا الحالي الولايات المتحدة الأميركية هي مثال ممتاز على الدولة الناجحة والمرنة، لأنها قائمة على اللامركزية والحرية.
الفكر المركزي التسلطي هو مخالف للطبيعة. لا يوجد في الطبيعة التي من حولنا أي شيء قائم على الإدارة المركزية المتسلطة. الطبيعة هي نظام معقد للغاية ولا يمكن لأية جهة مركزية أن تديرها، فما بالك بإنسان أحمق يظن أنه قادر على إدارة الطبيعة وفق هواه ونظرياته؟
لنعد إلى الموضوع الأصلي. أنا كنت أريد أن أقول أن الجهاز العصبي الحيواني هو من الأصل قائم على اللامركزية. لا يوجد في الجهاز العصبي للحيوانات منطقة مركزية وظيفتها الإشراف على بقية مناطق الجهاز العصبي. من يعتقد بوجود هكذا منطقة هو ببساطة لا يفهم الجهاز العصبي. لا يوجد في عالم الطبيعة شيء اسمه “إدارة مركزية”. هذه الفكرة هي مجرد خرافة بشرية. لا يوجد شيء في الطبيعة يعمل وفق هذه الآلية الخرافية. لو كان الجسم البشري يعمل وفق آلية الإدارة المركزية لما كان البشر استمروا ولكانوا انقرضوا منذ اليوم الأول لظهورهم.
الجسم الحيواني والبيئة المحيطة به هي أمور معقدة جدا ويحكمها عدد كبير من المتغيرات. الطريقة الوحيدة الممكنة لإدارة الجسم الحيواني هي الأسلوب غير المركزي الفضفاض، وهذا هو المطبق فعلا في الطبيعة.
الفهم الصحيح للجهاز العصبي هو أنه مجموعة من العناصر المتكتلة التي يؤازر بعضها بعضا. كل عنصر من هذه العناصر هو متخصص في وظيفة معينة، ولكن هذا لا يعني أن هناك عناصر “فوق” عناصر أخرى. لا يوجد في الجهاز العصبي ملوك وأمراء أو أحزاب اشتراكية تقدمية. هناك فقط أجزاء متخصصة تتعاون مع بعضها.
لهذا السبب عند الحديث عن الجهاز العصبي أنا لا أميل إلى استخدام مصطلح “اللوحة الأم” ونحو ذلك من المصطلحات المبنية على الثقافة البشرية. في الجهاز العصبي لا يوجد أم وأب. هناك في الجهاز العصبي أجزاء وظيفتها الربط association بين البيانات القادمة من مناطق مختلفة في الجهاز العصبي. الدماغ الأوسط midbrain) mesencephalon) هو أحد هذه الأجزاء، وهو ربما أول منطقة في الجهاز العصبي لعبت هذا الدور الهام. من الممكن أن الدور الأصلي للدماغ الأوسط لدى الفقاريات هو نفس دور الدماغ الثالث لدى الحشرات.
من الممكن أن الدماغ الأوسط لدى الفقاريات القديمة كان يلعب دورا في إنشاء السلوك. الدماغ الأوسط يحصل على البيانات البصرية والشمية من الدماغ الأمامي، وهو يحصل على البيانات الحسية القادمة من المناطق السفلى من الجهاز العصبي. بالتالي هو في موقع ممتاز لكي يربط البيانات ببعضها ويقرر تحفيز السلوك من عدمه. في الحقيقة الدماغ الأوسط ما يزال يؤدي هذا الدور عند البشر، ولكن الإشارات التي ينتجها الدماغ الأوسط عند البشر هي ذات طابع فضفاض. علماء النفس والجهاز العصبي يطلقون على هذه الإشارات مسمى “الحافز” motivation أو “الدافع” drive أو “الحاجة” need. هذه الكلمات هي وصف لوظيفة الدماغ الأوسط. الدماغ الأوسط هو الذي “يحفز” أو “يدفع” أو “يحيج” الإنسان للقيام بالسلوك (مهما كانت طبيعة السلوك). إذن دور الدماغ الأوسط هو محوري بالنسبة للسلوك البشري. بدون الحافز أو الدافع أو الحاجة فإن الإنسان سيتجمد ولن يتفاعل مع بيئته (وهو ما يحصل بالفعل في بعض الأمراض التي تصيب الدماغ الأوسط، من قبيل مرض باركنسون Parkinson’s disease). هذه الوظيفة التي يؤديها الدماغ الأوسط هي فضفاضة المعنى، ولكنها رغم ذلك محورية لعمل الجهاز العصبي. هذا إن دل على شيء فهو يدل على مدى بدائية هذه الوظيفة وقدمها.
على الأرجح أن وظيفة الدماغ الأوسط كانت محددة أكثر لدى الحيوانات الفقارية القديمة. في تلك الحيوانات الدماغ الأوسط كان ربما “ينشئ” أو “يبدأ” السلوك وليس “يحفز” السلوك. هناك فرق بين “البدء” بالسلوك و”تحفيز” السلوك. التحفيز يعني وجود جهة أخرى يجب أن تتخذ القرار النهائي. في الدماغ البشري هذه الجهة هي القشرة المخية. القشرة المخية تطورت لدى الثدييات على نحو كبير وظهرت فيها مناطق واسعة تؤدي وظيفة الربط association بين البيانات. هذا التطور في القشرة المخية همش دور الدماغ الأوسط وحوله من “منشئ” أو “بادئ” للسلوك إلى “محفز” أو “دافع” له.
بما أن الدماغ الأوسط كان ربما هو صاحب قرار السلوك في الحيوانات القديمة فهذا يفسر لماذا كان حجمه كبيرا في تلك الحيوانات وكان شكله يشبه شكل المخ في دماغ الثدييات. هو كان يؤدي نفس الوظيفة التي يؤديها المخ عند الثدييات: هو كان يقرر السلوك.
في مواضيع المدونة أنا أحرص على ذكر المصطلحات العلمية باللغة الإنكليزية لأن هذه اللغة حاليا هي لغة العلم بامتياز. من يريد أن يقرأ أو أن يبحث في أي موضوع علمي فهو سيحتاج حتما للمصطلح الإنكليزي. المصطلح الإنكليزي يمكن ترجمته بسهولة (عبر الإنترنت) إلى المصطلح الفرنسي أو الألماني أو الروسي إلخ. بالتالي أنا عندما أذكر المصطلح الإنكليزي فإنني أهدف إلى إفادة نفسي وإفادة قرائي.
النظام الحاكم في سورية يروج لبعض الكلمات العربية على أنها “مصطلحات” علمية، وهو يجبر الناس الواقعين تحت سيطرته على استخدام هذه “المصطلحات” وعدم استخدام المصطلحات الإنكليزية أو غيرها.
السؤال الذي يطرح نفسه هو هل “المصطلحات” التي يروج لها النظام السوري هي بالفعل مصطلحات؟
كلمة “مصطلح” هي مشتقة من “التصالح” بمعنى “الاتفاق”. عندما نقول أن كلمة معينة هي “مصطلح” فهذا يعني أن هذه الكلمة هي محل اتفاق وإجماع، ولكن يا ترى من هم الذين اتفقوا على استخدام مصطلحات النظام السوري؟
لا أحد في العالم يستخدم مصطلحات النظام السوري سوى جامعات النظام السوري. حتى الدول العربية لا تستخدم هذه المصطلحات.
النظام السوري لا يحكم سوى جزء ضئيل من متحدثي اللغة العربية. هو ليس في موقع يسمح له بفرض “المصطلحات” على العرب. على أرض الواقع معظم العرب لا يستخدمون مصطلحات النظام السوري ولكنهم يستخدمون المصطلحات المتداولة دوليا.
النظام السوري يتحدث منذ أكثر من خمسين سنة عما يسميه بـ”المصطلحات العلمية العربية”، ولكن عبارة “المصطلحات العلمية العربية” هي مجرد خرافة كبقية خرافات النظام السوري (وما أكثرها). “المصطلحات العلمية العربية” هي ليست سوى “مصطلحات سورية”. هذه المصطلحات هي غير مستخدمة خارج نطاق سلطة النظام السوري.
النظام السوري ليس معروفا بالمستوى العلمي العالي. سورية تحت سلطة النظام السوري هي من أكثر دول العالم تخلفا على الصعيد العلمي والبحثي. ما هي القيمة العلمية لمصطلحات النظام السوري؟ ما هي الفائدة من تعلم هذه المصطلحات؟
مصطلحات النظام السوري “العلمية” هي ليست أكثر من مسخرة على شاكلة مساخر العقيد الراحل القذافي (الذي كانت لديه أسماء أشهر خاصة به لا يستخدمها أحد غيره).
النظام السوري بسبب عقليته الجاهلة والخرقاء يعتقد أن “تعريب العلم” هو مجرد قرار حكومي لا يتطلب أكثر من توقيع رئيس الجمهورية.
اللغة الإنكليزية لم تصبح لغة العلم لأن الحكومة البريطانية اتخذت قرارا بذلك. اللغة الإنكليزية أصبحت لغة العلم بحكم الأمر الواقع. حاليا أهم الكتب والمؤلفات والأبحاث العلمية تصدر باللغة الإنكليزية. هذا هو السبب الذي أدى إلى “أنكلزة العلم”.
كلمة “تعريب العلم” هي كلمة ذات معنى ضخم للغاية. من يريد أن يعرّب العلم عليه أن يسعى لتطوير مستوى التعليم والبحث العلمي في الدول العربية. يجب أن يكون هناك إنتاج علمي غزير في الدول العربية وأن يكون العرب هم قادة البحث العلمي في العالم. عندما يصبح العالم العربي هو المصدر الأول للعلم يمكننا وقتها أن نتحدث عن “تعريب العلم”.
النظام السوري لا يفقه شيئا من ذلك. هو جالس في جورته العميقة ويقوم من هذه الجورة بإصدار الفرمانات وفرضها على المساكين الواقعين تحت سلطته، والذين هم قلة قليلة من سكان العالم.
هذه النزعة المرضية للتحكم والسيطرة هي سبب كل التخلف والمآسي التي عاشتها سورية.
أنا عندما أكتب فإنني أريد أن أفيد نفسي وأفيد الناس، وليس أن أفرغ أمراضي النفسية. أنا أريد لنفسي ولمن يتابعني أن نقرأ ونبحث ونفهم. هدفي ليس أن أشكّل الناس على مزاجي لكي يتحدثوا كما أريد ويفكروا كما أريد. أنا أصغر بكثير من أجعل الناس يتحدثون ويتصرفون على مزاجي. أنا أعرف حجمي وقدري في هذا العالم وأعرف أن ما أكتبه لا يساوي شيئا مقارنة بما يصدر في العالم من أبحاث وعلوم.
لو أنني استخدمت في مقالاتي “مصطلحات” النظام السوري فهذا لن يفيدني وقرائي بشيء، بل على العكس هو سيضرنا. مصلحتي ومصلحة القراء هي أن نعرف المصطلحات الدولية لكي نتمكن من البحث عن هذه المصطلحات في الإنترنت والكتب والدراسات. لو أنني ذكرت مصطلحات النظام السوري فما الذي سنفعله بهذه المصطلحات؟ هذه المصطلحات لا قيمة لها في العلم.
هي أصلا ليست مصطلحات، لأن لا أحد يستخدمها سوى جامعات النظام السوري. هي مجرد ترجمات وليست مصطلحات، وهي بصراحة ترجمات رديئة ولا فائدة من ذكرها. كثير من هذه الترجمات هي خاطئة (بمجرد الرجوع للقواميس اللاتينية واليونانية يتبين أن كثيرا من هذه الترجمات هي غير صحيحة أو غير دقيقة)، وكثير منها هي كلمات غير مألوفة لدى متحدثي اللغة العربية (بعضها كلمات عربية بائدة لا يستخدمها أحد في العصر الحالي، وبعضها كلمات لا أصل لها قام مترجمو النظام السوري باختراعها). لا توجد فائدة من ذكر هذه الترجمات لأنها هي نفسها تحتاج للترجمة. بالنسبة لي شخصيا فأنا لا أستطيع فهم معاني هذه الكلمات إلا بالرجوع إلى لسان العرب ونحوه. ما هي الفائدة من ذلك؟ لماذا أضيع وقتي ووقت القراء بالرجوع إلى قواميس اللغة العربية؟ ولماذا نملأ ذاكرتنا بكلمات لا فائدة من حفظها؟ الأجدى لي وللقراء هو أن نحفظ الكلمات الأجنبية المستخدمة دوليا وأن نرجع إلى القواميس الأجنبية لكي نفهم معاني هذه الكلمات.
طبعا لتحديد الأجدى لا بد أولا أن نحدد الهدف. أنا هدفي هو رفع مستوى العلم لدي ولدى قرائي. النظام السوري لديه هدف آخر مختلف تماما. النظام السوري يريد أن “يعرب العلم”. إذن هدف النظام يختلف عن هدفي. النظام السوري ليس مهتما بالعلم ولكنه يعتبر العلم حقلا للصراع الأيديولوجي السياسي. “مصطلحات” النظام السوري العلمية ليس المقصود منها خدمة العلم ولكن المقصود منها محاربة الإنكليز والأوروبيين وتحقيق انتصار ثقافي وهمي عليهم. لهذا السبب أنا لن أستخدم “مصطلحات” النظام السوري العلمية.
في مقالاتي لا توجد أية مصطلحات سوى المصطلحات الدولية (المصطلحات الدولية = الكلمات المتداولة على نطاق دولي). بالنسبة للكلمات العربية التي أستخدمها في مقالاتي فهي ليست “مصطلحات” ولكنها مجرد ترجمات. أنا لدي الحرية في الترجمة كما أشاء ولست مضطرا للتقيد بترجمات النظام السوري السيئة. الهدف من ترجماتي هو توضيح المعنى لنفسي وللقراء وليس التعقيد وشغل ذاكرتنا بكلمات لا توجد فائدة من حفظها.
تكوين الدماغ البشري معقد، وآلية عمله أعقد، ولكنني في هذا الموضوع سأحاول أن أبسط الأمور.
أنا افترضت في مقال سابق أن آلية عمل الدماغ من حيث المبدأ هي نفس آلية عمل الكومبيوتر، أي أن مكونات الدماغ الأساسية هي ما يلي:
أجهزة الإدخال والإخراج
الذاكرة
المعالج
بالإضافة إلى هذه المكونات هناك شيء مهم يجب أن نضيفه هو مصدر الطاقة power supply. مصدر الطاقة لدى أجهزة الكومبيوتر هو تيار الكهرباء. بالنسبة للدماغ البشري فالمسألة فيها بعض التعقيد، لأن الدماغ هو موجود داخل جسم حي، والدماغ نفسه مكون من خلايا حية.
جهاز الكومبيوتر هو مكون من قطع غير حية. هذه القطع لا تحتاج للطاقة لكي تستمر في الوجود. جهاز الكومبيوتر يحتاج الطاقة فقط لكي يعمل، أي أن الطاقة الكهربائية عمليا لا تغذي جهاز الكومبيوتر ولكنها تغذي الوظائف التي يقوم بها جهاز الكومبيوتر. من الممكن أن نفصل الطاقة كليا عن جهاز الكومبيوتر ورغم ذلك فإنه لن يتلف. كل ما سيحصل هو أن وظائفه ستتوقف.
الدماغ البشري هو مكون من خلايا حية (هي الخلايا العصبية التي تسمى “نيورونات” neurons). هذه الخلايا تحتاج للطاقة بشكل دائم فقط لكي تستمر في الوجود. لو أننا قطعنا الطاقة عن الدماغ فهو سيتلف خلال دقائق. إذن الدماغ البشري يحتاج الطاقة لكي يوجد وليس فقط لكي يعمل.
الكومبيوتر فيه دارة واحدة كبيرة تغذيه بالطاقة الكهربائية، وهذه الدارة تعطي عددا من الدارات الفرعية الأصغر. الدماغ البشري يعمل وفق مبدأ مشابه، ولكن الفرق هو أن الدماغ البشري يحوي عددا هائلا من الدارات الفرعية هو تقريبا 100 مليار دارة. هذا العدد هو عدد الخلايا العصبية المكونة للجهاز العصبي. كل خلية عصبية (وكل خلية حية) تحوي دارة أو دارات خاصة لإنتاج الطاقة اللازمة لعمل الخلية. سبب هذا النظام المعقد هو أن الكائنات الحية كانت في الأصل تتكون من خلية واحدةunicellular . لاحقا ظهرت الكائنات متعددة الخلايا multicellular عن طريق التكاثر الخلوي والالتصاق بين الخلايا الناتجة. إذن الكائن متعدد الخلايا هو في الأصل مجموعة من الكائنات الملتصقة مع بعضها والمتحدة وظيفيا. هذا المبدأ يشبه نوعا ما مبدأ التوائم السيامية.
فكرة الكائنات متعددة الخلايا تشبه من حيث المبدأ فكرة التوائم السيامية
مصدر طاقة الخلايا الحيوانية (الخلايا المكونة لأجسام الحيوانات) هو الطاقة الكيميائية المختزنة في جزيئات المركبات العضوية organic. مصدر المركبات العضوية هو الخلايا الحية (سواء النباتية أم الحيوانية أم غير ذلك). الحيوانات تعتمد في غذائها بشكل كامل على المركبات العضوية المكونة في أجسام الكائنات الحية الأخرى. الخلايا الحيوانية تقوم بتحطيم الجزيئات العضوية الكبيرة (السكريات والبروتينات والدهون) إلى جزيئات صغيرة (عملية التحطيم تسمى “تفاعلات الهدم” catabolism). عملية التحطيم تؤدي إلى تحرير الطاقة الكيمائية المختزنة في الجزيئات العضوية الكبيرة. هذه الطاقة تستخدم لتغذية تفاعلات كيميائية من أنواع أخرى (منها مثلا “التفاعلات البنائية” anabolism). إذن ما يحصل داخل الخلايا الحية هو أشبه بدورة كيميائية يتم خلالها تحطيم جزيئات عضوية كبيرة للحصول على الطاقة واستخدام هذه الطاقة لتغذية تفاعلات عضوية أخرى تفيد الخلايا الحية والكائن الحي. هذه الدورة هي عمليا “الحياة”. أي جسم يحوي في داخله دورة كيميائية كهذه الدورة ويملك القدرة على التكاثر هو “كائن حي”.
بالنسبة للخلايا العصبية فهي تعتمد بشكل أساسي على الجزيئات السكرية كمصدر للطاقة—تحديدا هي تعتمد على جزيئات سكر الغلوكوز glucose. جزيئات الغلوكوز تصل إلى الخلايا العصبية عبر الدم (الدورة الدموية). لو انقطع وصول جزيئات الغلوكوز إلى الخلايا العصبية لأي سبب (مثلا توقف الدورة الدموية) فهذا سيؤدي إلى تلف الخلايا العصبية وتوقفها عن العمل بشكل نهائي خلال دقائق قليلة. لا يمكن بعد ذلك إعادة الحياة إلى الخلايا العصبية أو تعويضها بخلايا أخرى (لأن الخلايا العصبية هي غير قابلة للتجدد عبر الانقسام، بخلاف خلايا الجلد مثلا).
أجهزة الإدخال والإخراج
أجهزة الإدخال المتصلة مع الدماغ هي “أعضاء الحس” sense organs. هذه الأعضاء توصل البيانات من خارج الدماغ إلى داخله. شعبيا هناك خمسة أعضاء حسية مشهورة هي العينان والأذنان والأنف والفم والجلد. هذا التقسيم هو ليس دقيقا من الناحية العلمية ولكنه يفيد في توضيح ما أقصده بأجهزة الإدخال input devices.
أعضاء الحس تتصل مع الدماغ بواسطة أعصاب nerves تسمى “الأعصاب الحسية” sensory nerves. الأعصاب هي أشبه بالأسلاك الكهربائية التي تصل أجهزة الإدخال مع الكومبيوتر.
أجهزة الإخراج توصل توصل البيانات من داخل الدماغ إلى خارجه. هناك الكثير من أجهزة الإخراج المتصلة مع الدماغ، ولكن جميع هذه الأجهزة يمكن اختصارها بعبارتين: أعضاء الحركة وأعضاء الإفراز (التي تفرز جزيئات كيميائية).
مثلا الجهاز التنفسي هو من أجهزة الإخراج لأن الدماغ يستخدمه لإنتاج الكلام (الكلام يخرج من الفم ولكن آلية إنتاجه تتطلب مشاركة جميع أعضاء الجهاز التنفسي). آلية إنتاج الكلام في الجهاز التنفسي تعتمد بشكل كامل على حركة الهواء وتقلص العضلات في السبيل التنفسي (السبيل الصوتي). حركة الهواء في السبيل التنفسي تحدث بسبب تقلص عضلات جدار الصدر والحجاب الحاجز diaphragm، وبالتالي المحصلة هي أن إنتاج الكلام في الجهاز التنفسي يحدث بسبب تقلص العضلات.
أيضا لغة الإشارة (التواصل عن طريق حركات اليدين وتعابير الوجه) تعتمد على تقلص العضلات.
الدماغ يقوم بتقليص العضلات عن طريق أعصاب تسمى “الأعصاب المحركة” motor nerves.
بالإضافة إلى تقليص العضلات الدماغ يقوم أيضا بإخراج البيانات أو الرسائل على شكل جزيئات كيميائية. هذه الجزيئات تسمى “نواقل عصبية” neurotransmitters. النواقل العصبية تصنع داخل الخلايا العصبية وهي تستخدم داخل الدماغ نفسه في نقل الإشارات من خلية عصبية إلى خلية عصبية أخرى. الدماغ يستخدم هذه النواقل أيضا للتأثير في أعضاء الجسم الأخرى. الأعصاب التي تنقل النواقل العصبية إلى خارج الجهاز العصبي تسمى أحيانا “الأعصاب الإفرازية” secretory nerves.
الدماغ أيضا يقوم بتصنيع بعض الهرمونات hormones وطرحها في الدم بشكل مباشر (عن طريق الفص الخلفي من الغدة النخامية pituitary gland). الهرمونات هي عبارة عن جزيئات عضوية معقدة (بعضها ذات طبيعة بروتينية وبعضها ذات طبيعة دهنية) يتم إفرازها من أعضاء تسمى الغدد glands وتنتقل عبر الدم وتحدث تأثيرات في أعضاء الجسم.
لو درسنا جميع وسائل الإخراج التي يستخدمها الدماغ فسنجد أنها في المحصلة تعتمد على تقلص العضلات أو على إفراز النواقل الكيميائية. إشارات تقلص العضلات تنقل عبر الأعصاب المحركة، وإشارات الإفراز تنقل عبر الأعصاب الإفرازية.
مجموع الأعصاب الحسية والمحركة والإفرازية يسمى “الجهاز العصبي المحيطي” peripheral nervous system. هذه التسمية هي عمليا مجرد مرادف لكلمة “أعصاب”.
الجهاز العصبي المركزي
“الجهاز العصبي المركزي” central nervous system هي تسمية تشمل كلا من الدماغ brain والحبل الشوكي spinal cord (الحبل الشوكي يسمى في معظم لغات العالم “النخاع الشوكي”، ولكن هذه التسمية نادرة الاستخدام في الإنكليزية).
الجهاز العصبي المركزي هو معقد التكوين، والسبب هو أن هذا الجهاز لم يتكون خلال يوم وليلة ولكنه تطور تدريجيا خلال مئات ملايين السنين. الجهاز العصبي المركزي هو أعقد شيء في جسم الإنسان، وهذا التعقيد هو الميزة الأساسية التي تميز الإنسان عن بقية الحيوانات.
لتسهيل فهم الجهاز العصبي المركزي يجب في رأيي أن نفهم تاريخ هذا الجهاز وكيف تطور خلال مئات ملايين السنين. الجهاز العصبي هو مكون من أنسجة عصبية (النسيج tissue هو عبارة عن عدد كبير من الخلايا المتشابهة الملتصقة مع بعضها والتي تؤدي وظيفة واحدة). النسيج العصبي مكون من خلايا عصبية (نيورونات neurons). الخلية العصبية هي خلية متخصصة في نقل الإشارات العصبية.
أنواع الأنسجة الخلوية لدى الحيوانات الفقارية vertebrates (ومنها الإنسان)
كل الخلايا الحية التي تحوي نواة nucleus (الخلايا “حقيقية النواة” eukaryote) هي مشحونة كهربائيا. السبب هو وجود أنزيمات في أغشية هذه الخلايا تعمل على نقل بعض الأيونات ions (الجزيئات المشحونة كهربائيا) من داخل الخلية إلى خارجها (من السائل داخل الخلوي intracellular fluid إلى السائل خارج الخلوي extracellular fluid). هذا النوع من الأنزيمات يسمى “مضخات الأيونات” ion pumps. هذه المضخات ظهرت لأول مرة مع ظهور الخلايا ذات النواة eukaryotes قبل حوالي 1500 مليون عام.
في الأحوال الطبيعية (مثلا في الخلايا البدائية التي لا تملك نواة prokaryotes) الشحنة الكهربائية الإجمالية للسائل داخل الخلوي يجب أن تكون مساوية للشحنة الكهربائية الإجمالية للسائل خارج الخلوي، لأن الانتشار الجزيئي molecular diffusion يعمل على تحقيق التوازن الكهربائي-الكيميائي electrochemical equilibrium على جانبي غشاء الخلية. مضخات الأيونات تعمل على تخريب هذا التوازن. مثلا مضخات أيون الكالسيوم Ca2+ (التي يعتقد أنها أقدم أنواع المضخات الأيونية) تعمل على نقل أيون الكالسيوم من داخل الخلية إلى خارجها بشكل أعمى وبغض النظر عن تركيزه في الداخل والخارج. هذه المضخات لا تتأثر بالانتشار الجزيئي بل هي تقاومه وتعمل ضده. هي تستفيد من الطاقة الكيميائية الناتجة من تفاعلات الهدم catabolism وتسخر هذه الطاقة لمقاومة الانتشار الجزيئي وخلق وضع غير متوازن على جانبي غشاء الخلية.
مقطع عرضي في غشاء خلوي تظهر فيه مضخات أيونية (بروتينات ناقلة carrier proteins)
بعد مضخات أيون الكالسيوم ظهرت أنواع أخرى من المضخات تضخ أيون البوتاسيوم K+. مضخات أيوني الكالسيوم والبوتاسيوم تهيمن لدى الحيوانات المكونة من خلية واحدة (“الحيوانات البدائية” protozoans). الحيوانات الأكثر تعقيدا تعتمد بشكل أكبر على مضخات أيون الصوديوم Na+ (المهمة لعمل الخلايا العصبية). مضخات أيون الصوديوم ظهرت لأول مرة لدى مجموعة من الحيوانات تسمى “الحيوانات الخيطية” filozoans. هذه الحيوانات كانت في الأصل وحيدة الخلية unicellular (مكونة من خلية واحدة)، ولكن أحد فروعها تطور قبل أكثر من 650 مليون عام وأصبح متعدد الخلايا multicellular (أي أنه يصبح يتكون من أنسجة tissues). هذا الفرع متعدد الخلايا يسمى “الحيوانات” animals (باللاتينية animalia)، ويسمى أيضا “الحيوانات التالية” metazoans. الحيوانات animals هي حسب علمي أول مجموعة ظهرت لديها الخلايا العصبية بالمعنى الحقيقي للكلمة.
تطور الجهاز العصبي لدى الحيوانات animals هو مرتبط بمدى اعتمادها على مضخات أيون الصوديوم. مثلا الحيوانات المسماة “الإسفنجيات” poriferans (والتي هي من أكثر أنواع الحيوانات بدائية) تعتمد بشكل كبير على مضخات أيون الكالسيوم، وهي لا تملك جهازا عصبيا حقيقيا. الحيوانات المسماة “المِشْطِيات” ctenophores و”اللاسعات” cnidarians تعتمد بشكل أكبر على مضخات أيون الصوديوم، وهي تملك جهازا عصبيا، ولكن هذا الجهاز هو مكون من نسيج عصبي بدائي يسمى “شبكة عصبية” nerve net. الشبكة العصبية هي عبارة عن مجموعة من الخلايا العصبية المتصلة مع بعضها على شكل شبكة، ولكنها لا تحوي دماغا أو أية منطقة مركزية وظيفتها الإشراف على عمل الشبكة.
المرحلة التالية في تطور الحيوانات كانت ظهور الحيوانات “ثنائية الجانب” bilaterians. أجسام هذه الحيوانات تتميز بوجود رأس وفم وشرج، وداخل الجسم يوجد نسيج عصبي ممتد على طول الجسم على شكل حبل يسمى “الحبل العصبي” nerve cord. من هذا الحبل تخرج الأعصاب نحو بقية مناطق الجسم.
تصوران افتراضيان لأول حيوان ثنائي الجانب urbilaterian
الحيوانات الحبلية chordates هي فرع من ثنائيات الجانب bilaterians. في هذه الحيوانات حصل تطور مهم يتعلق بتوزع مضخات الصوديوم في الخلايا العصبية (مضخات الصوديوم صارت تتركز في جانب واحد من الخلية العصبية). هذا التطور ترافق مع بروز نزعة نحو “الرأسنة” cephalization (مشتقة من كلمة “رأس” cephalē). المقصود بالرأسنة هو أن الحبل العصبي (الذي له شكل أنبوب) صار يميل إلى الانتفاخ في جهة واحدة من الجسم (هي الرأس). هذا الانتفاخ هو أساس فكرة “الدماغ” الذي ظهر لاحقا.
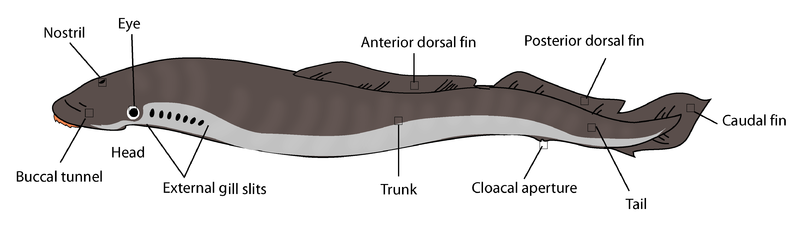
تصور افتراضي لجسم حيوان حبلي بدائي. كلمة gut تعني “أمعاء”، وكلمة anus تعني “شرج”مخطط لجسم حيوان حبلي بدائي يعيش حاليا اسمه lancelet أو amphioxus (باللاتينية Branchiostoma). الحبل العصبي nerve cord يظهر باللون الأصفر. الرقم 1 يشير إلى الانتفاخ الدماغي في منطقة الرأس، والرقم 17 يشير إلى الأعصاب. الرقم 2 يشير إلى الحبل الظهري notochord (الذي هو دعامة الجسم بدلا من العمود الفقري الموجود لدى الفقاريات). جهاز الدوران الدموي يظهر باللون الأحمر. الرقم 15 يشير إلى الغدد التناسلية (المبايض أو الخصى). الرقم 19 يشير إلى الكبد. العضو ذو اللون الأخضر هو الفم. المناطق ذات اللون البني-الأخضر هي الجهاز الهضمي (يبدأ من الفم وينتهي في الشرج). على ما يبدو فإن جهاز الإخراج البولي لا يظهر على هذا الرسم.
ما يلي صور أخرى لنفس الحيوان lancelet (هذا الحيوان ينتمي للحبليات الرأسية cephalochordates التي هي شكل بدائي من الحبليات ما زال يعيش اليوم في معظم بحار العالم):
كلمة lancelet هي تصغير لـ lancet التي تعني “مبضع جراحي”.
الحيوانات الفقارية vertebrates تطورت من حيوان حبلي بدائي شبيه بـ lancelet. هذا التطور حصل قبل أكثر من 500 مليون عام. الفرق الأساسي بين الفقاريات والحبليات غير الفقارية هو أن الفقاريات تملك عمودا فقاريا vertebral column يشكل دعامة عظمية للظهر بدلا من الحبل الظهري notochord (بقايا الحبل الظهري تشكل لدى الفقاريات ما يسمى “النواة اللبية” nucleus pulposus التي توجد داخل “الأقراص بين الفقارية” intervertebral disc).
مقطع في العمود الفقري تظهر فيه “النواة اللبية” nucleus pulposus باللون البنفسجي. في هذا الرسم النواة اللبية هي في حالة فتق (متمددة إلى خارج مكانها الطبيعي) وهي تضغط على العصب المتصل مع الحبل الشوكي. هذه الحالة المرضية تسمى “فتق النواة اللبية” nucleus pulposus herniation أو “فتق القرص الشوكي” spinal disc herniation، وهي نفس الحالة التي يسميها الناس “مرض الديسك”
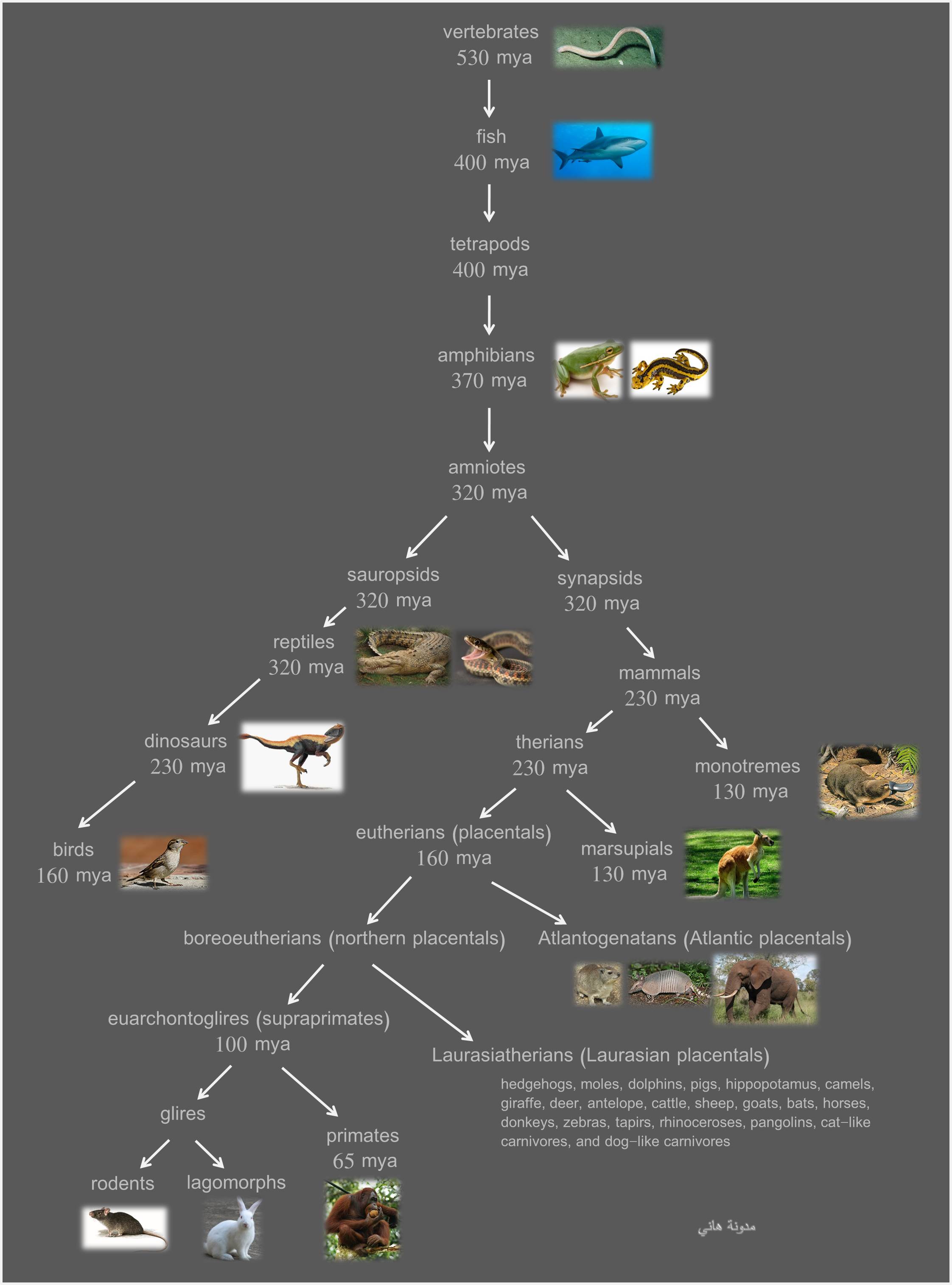
الفقاريات تصنف تقليديا ضمن خمس مجموعات:
الأسماك fish
البرمائيات amphibians
الزواحف reptiles
الطيور birds
الثدييات mammals
هذا التصنيف هو تصاعدي من الأقل تطورا نحو الأكثر تطورا. الأسماك هي أقل الفقاريات تطورا، والثدييات هي أكثر الفقاريات تطورا.
هذا التصنيف هو مجرد تصنيف شعبي وليس تصنيفا علميا، بمعنى أنه لا يعبر عن شجرة تطورية صحيحة. الباحثون كانوا يعلمون ذلك منذ زمن بعيد وكانوا يحاولون أن يرسموا شجرة تطورية علمية (phylogenetic)، وهم تمكنوا من تحقيق تقدم كبير مؤخرا بسبب الدراسات الجزيئية والجينية، ولكن المشكلة هي أن الأشجار التي يرسمونها هي معقدة للغاية. أنا لن أنقل الأشجار التي يرسمونها ولكنني سأعرض شجرة مبسطة:
هذه الشجرة هي مختصرة جدا ولكنها رغم ذلك مفيدة لفهم السياق العام لتطور الفقاريات. الفقاريات البدائية كانت كلها تعيش في الماء (كل أو معظم الحيوانات البدائية كانت تعيش في الماء). كلمة “الأسماك” fish هي ليست تصنيفا علميا صحيحا لأن هذه الكلمة تشمل أنواعا مختلفة من الفقاريات البدائية التي تطورت بشكل مستقل وفي أزمنة مختلفة. ما يسمى بـ “الأسماك” هو عمليا كل الفقاريات التي تعيش في الماء.
أكثر الفقاريات بدائية هي على ما أظن الكائنات التي تسمى “الأسماك بلا فكين” jawless fish. هذه التسمية هي ليست تصنيفا علميا ولكنها تسمية تشمل بعض الفقاريات البدائية للغاية من قبيل الحيوان المسمى lamprey. هذا الحيوان هو طفيلي يعيش في البحر ويتغذى من امتصاص دماء الأسماك (وفي بعض الأحيان يتسبب بنفوق أعداد كبيرة منها).
lampreyالـ lamprey له رأس وعينان ولكنه لا يملك فكينسمكة ضحية للـ lamprey
هناك حيوان شبيه بالـ lamprey ولكنه أكثر بدائية منه اسمه hagfish. هذا الحيوان لا يملك عمودا فقريا حقيقيا، ولهذا السبب بعض الباحثين يرفضون تصنيفه ضمن الفقاريات.
hagfish
الأسماك بلا فكين كـ lamprey و hagfish هي ربما أكثر الكائنات المعاصرة شبها بالفقاريات الأولى التي ظهرت قبل حوالي 500 مليون عام.
الأسماك الغضروفية cartilaginous fish تأتي في المرتبة الثانية من حيث التطور بعد الأسماك بلا فكين. هذه الكائنات تملك هكيلا غضروفيا بدلا من الهيكل العظمي، وأبرز مثال عليها هو سمك القرش.
سمك القرش
بعد الأسماك الغضروفية ظهرت الأسماك العظمية bony fish التي تشمل معظم الأسماك المعروفة حاليا.
قبل 400 مليون عام حدثت طفرة كبيرة في تطور الفقاريات أدت إلى ظهور الفقاريات “رباعية الأرجل” tetrapods. هذه الكائنات امتلكت لأول مرة الأطراف الأربعة (الذراعان والرجلان). الفقاريات التي كانت تعيش قبل ذلك كانت تملك رأسا وجسما ولكنها لم تكن تملك ذراعين ورجلين.
مجموعة من رباعيات الأرجل الباكرة. الصورة مبنية على مستحاثات
رباعيات الأرجل هي أولى الفقاريات التي خرجت من الماء إلى البر. البرمائيات amphibians (كالضفادع والسلمندر) هي ربما الأكثر بدائية من بين رباعيات الأرجل الموجودة حاليا، بمعنى أنها الأكثر شبها برباعيات الأرجل الأولى. البرمائيات تستطيع أن تعيش خارج الماء بعد أن يكتمل نموها، ولكنها لا تستطيع أن تعيش خارج الماء في مراحل حياتها الباكرة (المرحلة الجنينية). لهذا السبب البرمائيات تضع بيوضها في الماء أو في الأماكن الرطبة.
الحيوانات السلوية amniotes هي شكل متطور من رباعيات الأرجل. هذه الحيوانات لا تحتاج لأن تعيش في الماء مطلقا ولا حتى في المرحلة الجنينية. لهذا السبب هي تضع بيوضها خارج الماء وفي أماكن جافة. سر هذه الحيوانات هو أن أطفالها يحاطون في المرحلة الجنينية بغشاء يسمى السَّلَى amnion. هذا الغشاء يشكل حول الجنين كيسا مملوءا بالماء اسمه “الكيس السلوي” amniotic sac (يسمى أيضا “كيس الماء” water sac).
جنين بشري داخل الكيس السلوي في الرحم
الكيس السلوي هو أشبه “ببحر صناعي” يحيط بجنين الحيوانات السلوية ويغنيه عن العيش في البحر الحقيقي.
الحيوانات السلوية انقسمت قبل حوالي 300 مليون عام إلى فرعين كبيرين، فرع يسمى sauropsids (“سحليات الشكل”) وفرع يسمى synapsids (“ملتحِمات القوس”) أو theropsids (“وحشيات الشكل”). الفرع الأول أنتج الزواحف reptiles التي هيمنت على العالم طوال الحقبة المسماة Mesozoic Era (“حقبة الحياة الوسطى”). هذه الحقبة تسمى أيضا “عصر الزواحف” Age of Reptiles. خلال هذه الحقبة كانت توجد زواحف عملاقة هي الديناصوراتdinosaurs (معنى الكلمة هو “السحالي الفظيعة” terrible lizards). الديناصورات تعرضت لكارثة أدت إلى انقراض معظمها قبل 66 مليون عام. الطيور birds هي الفرع الوحيد المتبقي من الديناصورات، ولكن معظم الطيور المعاصرة هي صغيرة الحجم.
تطور الطيور من الديناصورات
الحيوانات المسماة synapsids كانت معاصرة للزواحف الباكرة ولكنها كانت مهمشة وصغيرة الحجم. الثدييات mammals هي فرع من هذه الحيوانات. الميزة الأساسية للثدييات هي غدة الثدي mammary gland التي تفرز الحليب. الثدييات عموما تلد ولا تبيض. الاستثناء الوحيد هو فرع باكر من الثدييات يسمى monotremes. الثدييات التي تنتمي لهذا الفرع تعيش في أستراليا وهي تشمل مثلا الحيوان المسمى platypus.
platypus
بقية الثدييات تسمىtherians (“الوحوش”). هذه الكائنات ظهرت قبل حوالي 200 مليون عام وهي تتميز عن كل الحيوانات الأخرى بأنها تلد بدل أن تبيض. الفرع المسمى marsupials (“الجيبيات”) له عدد من الخصائص الغريبة التي تميزه عن بقية الثدييات: إناث الحيوانات المنتمية لهذا الفرع يملكن رحما مقسوما إلى حجرتين، وهناك مهبل خاص يوصل إلى كل حجرة من الحجرتين، أي أن الأنثى تملك مهبلين. الذكر يملك قضيبا متفرعا إلى فرعين. عند الجماع الذكر يدخل فرعي قضيبه في مهبلي الأنثى. عندما تحمل الأنثى فإنها تلد الطفل بسرعة بالغة (خلال خمسة أسابيع)، والسبب هو أن الكيس السلوي لا يتثبت على جدار الرحم بواسطة مَشِيمة placenta. المشيمة هي عضو هام يربط الكيس السلوي بالجدار الداخلي للرحم، ومن خلال المشيمة يتم تزويد محتويات الكيس السلوي (خاصة الجنين) بالغذاء والأكسجين، ويتم أيضا تخليصها من الفضلات. غياب المشيمة عند إناث الحيوانات الجيبية يعني أن الجنين لا يمكنه أن يظل في الرحم لفترة طويلة وإلا فإنه سيموت. لهذا السبب الجنين يخرج من الرحم بسرعة بالغة ويعيش داخل جيب على بطن الأم إلى أن يكتمل نموه.
أشهر مثال على الحيوانات الجيبية هو ربما الكنغرو kangaroo
بقية الثدييات تسمىeutherians (“الوحوش الحقيقية”) أو placentals (“المشيميات”). هناك فروع عديدة جدا للمشيميات لا يمكن أن نخوض فيها الآن. أهم فرع من المشيميات بالنسبة لموضوعنا هو الرئيسيات primates (تحدثت عنه في مقال سابق). أقرب فروع المشيميات إلى الرئيسيات يسمى glires. هذا الفرع يضم كلا من الأرانب والقوارض (القوارض تشمل الفئران والجرذان والسناجب). الأصل المشترك للرئيسيات والأرانب والقوارض عاش قبل 100 مليون عام تقريبا.
الثدييات كانت مهمشة في عصر الزواحف Mesozoic Era، ولكن بعد الكارثة التي أصابت الديناصورات قبل 66 مليون عام بدأت حقبة جديدة تسمى Cenozoic Era (“حقبة الحياة الجديدة”). هذه الحقبة تسمى أيضا “عصر الثدييات” Age of Mammals لأن انقراض الزواحف الكبيرة أفسح المجال أمام الثدييات لكي تهيمن على العالم خلال هذه الحقبة.
حقبة Cenozoic Era ما زالت مستمرة حتى الآن ولم تنته بعد.
الإشارة العصبية
ذكرنا في الأعلى أن الخلايا الحية التي تملك نواة (“حقيقية النواة” eukaryotic) هي مشحونة كهربائيا. السبب هو وجود مضخات أيونية ion pumps في أغشية هذه الخلايا تستغل الطاقة الكيمائية الناتجة من تفاعلات الهدم catabolism لكي تتلاعب بتراكيز الأيونات في السائل داخل الخلوي intracellular fluid.
أهم أنواع المضخات الأيونية في خلايا الثدييات هي المضخة المسماة Na+/K+ ATPase. هذه المضخة تستغل الطاقة الكيميائية المختزنة في جزيء (adenosine triphosphate) ATP لكي تنقل ثلاثة أيونات من الصوديوم إلى خارج الخلية في مقابل نقل أيونين من البوتاسيوم إلى داخل الخلية. المحصلة الكيميائية لعمل هذه المضخة هي تقليل تركيز أيونات الصوديوم وزيادة تركيز أيونات البوتاسيوم داخل الخلية، والمحصلة الكهربائية هي خلق شحنة سلبية داخل الخلية (لأن المضخة تخرج ثلاثة أيونات صوديوم يحمل كل منها الشحنة 1+ وتدخل أيوني بوتاسيوم يحمل كل منهما الشحنة 1+، إذن المحصلة الكهربائية لعمل المضخة هي إخراج شحنة مقدارها 1+ في كل مرة تعمل فيها).
صورة توضح عمل مضخة Na+/K+ ATPase في الغشاء الخلوي
بالإضافة إلى المضخة السابقة هناك مضخات تنقل أيون الكالسيوم Ca2+ إلى خارج الخلية. هناك مضخة Ca2+ ATPase التي تستغل الطاقة الكيميائية المختزنة في جزيء ATP لكي تنقل أيونا واحدا من الكالسيوم إلى خارج الخلية. هذه المضخة هي ربما أقدم أنواع المضخات الأيونية، ولكن مشكلتها أن عملها بطيء ولا يفيد في الأنسجة التي تتطلب إخراج كميات كبيرة من أيون الكالسيوم في وقت قصير (مثلا الأنسجة العصبية والعضلية). لهذا السبب هناك مضخة أخرى أسرع من هذه المضخة هي Na+/Ca2+ exchanger. هذه المضخة لا تستهلك الطاقة الكيميائية ولكنها تستفيد من الفارق في تركيز الصوديوم بين السائل خارج الخلوي والسائل داخل الخلوي لتوليد الطاقة. هي تسمح لثلاثة أيونات صوديوم بالدخول إلى الخلية في مقابل إخراج أيون كالسيوم واحد. هذا الأمر لا يؤثر على تركيز الصوديوم داخل الخلية سوى بشكل مؤقت، لأن هذه المضخة لا تعمل أصلا إلا في مناسبات معينة (عندما يكون هناك تركيز عال من الكالسيوم داخل الخلية، كما يحصل مثلا عند تقلص الخلايا العضلية).
الرسم التالي يلخص أنواع المضخات التي ذكرناها:
المحصلة النهائية لعمل المضخات التي ذكرناها هي أن تركيز الصوديوم والكالسيوم يكون منخفضا داخل الخلية مقارنة بالسائل خارج الخلوي، وتركيز البوتاسيوم يكون مرتفعا داخل الخلية مقارنة بالسائل خارج الخلوي. هناك أيضا شحنة كهربائية سلبية تتولد داخل الخلية. هذه الشحنة تؤدي إلى طرد الأيونات ذات الشحنة السلبية إلى خارج الخلية، خاصة أيون الكلور Cl− الذي ينخفض تركيزه داخل الخلية إلى رقم ضئيل جدا. خروج أيون الكلور من الخلية لا يفيد في تعديل الشحنة الكهربائية لأن الخلية تحوي في داخلها الكثير من الجزيئات العضوية الثقيلة ذات الشحنة السلبية (خاصة الأحماض الأمينية amino acids المكونة للبروتينات). هذه الجزيئات العضوية الثقيلة لا يمكنها أن تخرج عبر الغشاء الخلوي، وبالتالي عمليا لا يوجد حل لتعديل الشحنة السلبية سوى بإدخال جزيئات ذات شحنة إيجابية من الخارج، وهذا الأمر لا يحدث في الأحوال الطبيعية.
لهذا السبب الخلايا في الأحوال العادية هي مشحونة سلبيا. الشحنة السلبية تؤدي إلى توليد مجال كهربائي بين السائل داخل الخلوي والسائل خارج الخلوي. هذا المجال الكهربائي يولد ما يسمى في الفيزياء باسم “فرق الكمون الكهربائي” electric potential difference أو الفولتاج voltage. قيمة فرق الكمون الكهربائي داخل الخلايا البشرية تتراوح في الأحوال العادية بين 60- و 80- ميلي فولت. فرق الكمون الكهربائي يترافق مع فرق كمون كيميائي ناجم عن الفرق الكبير في تراكيز الأيونات بين السائل خارج الخلوي والسائل داخل الخلوي. محصلة الكمونين الكهربائي والكيميائي تسمى “الكمون الكهربائي-الكيميائي” electrochemical potential.
في البيولوجيا الكمون الكهربائي-الكيميائي للخلايا في الأحوال العادية يسمى “كمون الراحة” resting potential.
الخلايا العصبية تستفيد من هذا الكمون لتوليد الإشارات العصبية.
ما يحصل باختصار هو أن الخلايا العصبية تسمح لأيونات الصوديوم بالدخول إليها من جهة واحدة. بما أن الخلية العصبية هي طويلة جدا (في بعض الأحيان طولها يمكن أن يصل إلى متر كامل لدى البشر) فهذا يؤدي إلى خلق فرق في الكمون داخل الخلية نفسها: في أحد طرفي الخلية هناك ارتفاع سريع في قيمة الكمون بسبب دخول أيون الصوديوم بكميات كبيرة، وفي الطرف الآخر من الخلية الكمون ما زال على حاله. هذا الفرق في الكمون داخل الخلية يؤدي إلى انتشار كهربائي-كيميائي من أحد طرفيها إلى الطرف الآخر. هذا باختصار شديد هو مبدأ الإشارة العصبية. الإشارة العصبية هي موجة كهربائية-كيميائية (مكونة أساسا من أيونات الصوديوم Na+) تنتقل من أحد طرفي الخلية العصبية إلى طرفها الآخر.
هذا الرسم من ويكيبيديا يوضح الفكرة:
الإشارات الصفراء تعبر عن أيونات الصوديوم. انتشار أيونات الصوديوم على الجانب الداخلي من الغشاء الخلوي يحفز دخول المزيد من أيونات الصوديوم من الخارج
البروتينات الناقلة في الغشاء الخلوي تقوم بإعادة الكمون السلبي إلى داخل الخلية بسرعة بالغة. لهذا السبب نلاحظ على الرسم أن الكمون الإيجابي المنتشر داخل الخلية يختفي بسرعة. في المحصلة الإشارة العصبية هي مجرد نبضة.
التغيرات الكهربائية-الكيميائية التي تحدث داخل الخلية أثناء مرور الإشارة العصبية تسمى “كمون الفعل” action potential.
كمون الفعل ليس خاصا بالخلايا العصبية ولكنه يحصل أيضا في الخلايا العضلية (ويؤدي إلى تقلصها). طريقة كمون الفعل في الخلايا العضلية تختلف عنه في الخلايا العصبية. في الخلايا العضلية ليست هناك حاجة لنقل إشارة من أحد طرفي الخلية إلى طرفها الآخر. لهذا السبب كمون الفعل في الخلايا العضلية لا يبدأ من طرف الخلية ولكنه يبدأ من وسطها أو من أي مكان فيها، وهو ينتشر في كل الاتجاهات. في الخلايا العضلية كمون الفعل يؤدي إلى تدفق أيون الكالسيومCa2+ إلى داخل الخلايا، وهذا تحديدا هو سبب تقلص اللييفات العضلية myofibrils.
كمون الفعل في الخلايا العضلية
طريقة كمون الفعل في الخلايا العضلية هي أكثر بدائية من طريقته في الخلايا العصبية، لأن الخلايا العصبية هي اختراع أحدث من الخلايا العضلية. الخلايا العصبية ظهرت لأول مرة مع ظهور الحيوانات متعددة الخلايا، والنقل العصبي بالطريقة الموجودة لدى البشر بدأ يظهر لأول مرة مع ظهور الحبليات chordates. النقل العصبي تطور لدى الحبليات بشكل متدرج. أحد التطورات المهمة التي حدثت لدى الحبليات الباكرة هو تركيز مضخات الصوديوم في منطقة معينة من الخلية العصبية، وبعد ذلك ظهر غمد الـ myelin الذي زاد من سرعة النقل في الخلايا العصبية بشكل كبير.
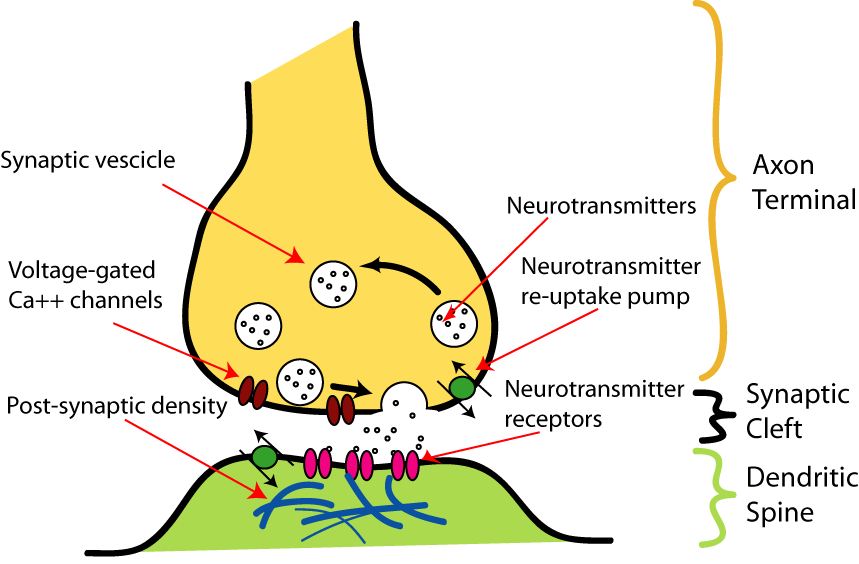
الخلية العصبية بشكلها الحديث تتألف من “جسم” body يحوي “شَجَرِيات” dendrites ومن استطالة طويلة تسمى “المحور” axon. المحور في الخلايا ذات النقل السريع يكون مغطى بغمد sheath من مادة اسمها myelin. الخلايا التي تنتج هذا الغمد في الجهاز العصبي المحيطي اسمها خلايا Schwann، وفي الجهاز العصبي المركزي اسمها “الخلايا قليلة الشجريات” oligodendroctyesمناطق التشابك بين الخلايا العصبية والخلايا الأخرى تسمى synapsesتشابك محور عصبي مع ليف عضلي (الليف العضلي هو مجموعة من الخلايا العضلية الملتحمة مع بعضها طوليا)نقل الإشارة عبر منطقة التشابك العصبي synapse يتم بواسطة جزيئات كيميائية تسمى “نواقل عصبية” neurotransmitters. هذه الجزيئات تتحرر من حويصلات في منطقة التشابك العصبي بسبب وصول كمون الفعل إليها
أجسام الحيوانات طورت مبدأ النقل العصبي بشكل تدريجي، وهذا ما منحها القدرة على نقل البيانات بسرعة فائقة. لاحقا الجهاز العصبي تطور لدى الثدييات على نحو غير مسبوق، وخاصة لدى الرئيسيات، وبالأخص لدى البشر.




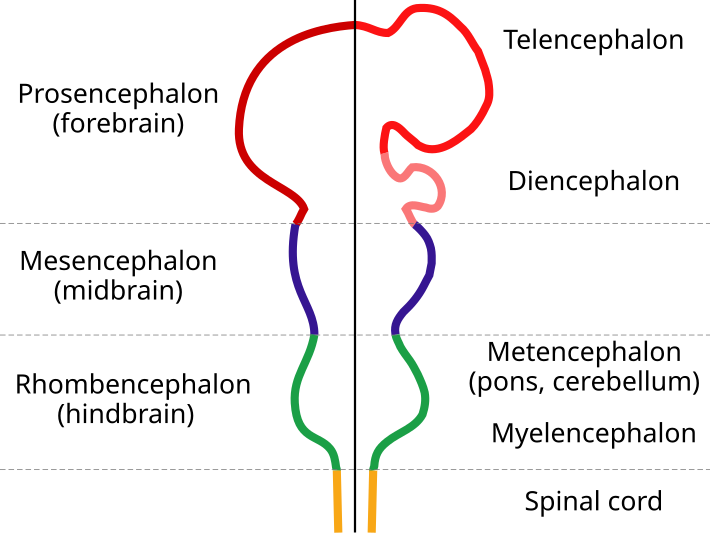







.jpg)






























































{kind=link}
{kind=link}